Une transpiration nécessaire
La transpiration des plantes est élevée et peut atteindre deux fois le poids de la plante un jour d’été, contre 2 à 4 % du poids chez un animal (voir Chap. II.5 et II.6). Le besoin en eau est imposé aux plantes par les conditions climatiques (rayonnement solaire, humidité relative de l’air, vitesse du vent, voir Chap. II.5). Il dépend de plus de la surface de feuilles par unité de surface du sol, et devient maximal quand chaque m² de sol est couvert par plus de 4 m² de feuilles. Les différences de transpiration entre espèces passent par ces deux sources de variation que sont le climat et la surface foliaire. Si 1 m² de maïs transpire plus par jour qu’1 m² de blé, c’est parce qu’il croît en été avec un rayonnement et une sécheresse de l’air plus forts que pendant la croissance du blé au printemps. Il n’y a donc rien à attendre de la génétique (sélection ou OGM) pour réduire les besoins des plantes en transpiration.
Pourquoi les plantes ont elles évolué vers des systèmes associés avec d’aussi fortes transpirations (voir figure ci-dessous) ? Premièrement, la transpiration permet à la plante de ne pas s’échauffer au soleil, comme le fait une voiture en stationnement. L’évaporation de l’eau rafraîchit les feuilles et les maintient ainsi à une température compatible avec leur métabolisme (voir Chap. II.3 et IV.1). Deuxièmement, la transpiration est nécessaire à la croissance et à la photosynthèse. Le gaz carbonique (CO2) pénètre dans les feuilles par des pores microscopiques, les stomates. C’est par ces mêmes stomates que l’eau est transpirée par la plante, si bien que toute réduction de transpiration s’accompagne d’une réduction de photosynthèse. Il n’y a aucun moyen connu d’éviter cet échange « eau contre carbone ». Cependant, le rapport entre photosynthèse et transpiration (efficience de l’eau transpirée, voir Chap. II.10) n’est pas constant, si bien qu’il peut être manipulé génétiquement. Par exemple, une variété de blé a été développée en Australie, qui a une meilleure efficience et des rendements augmentés de 10 % dans les conditions très sèches du sud australien (Rebetzke et al., 2002).
Que se passe-t-il si la plante ne peut pas prélever la quantité d’eau dont elle a besoin en fonction du climat ? D’abord, elle ralentit sa transpiration en fermant partiellement ou totalement ses stomates. Cela évite la déshydratation des feuilles, mais cause leur échauffement et réduit la photosynthèse. À plus long terme, la plante réduit sa surface foliaire, si bien que sa transpiration est réduite mais la photosynthèse également. D’autres adaptations existent, qui dépendent des espèces considérées : l’enroulement ou l’orientation des feuilles, un développement racinaire plus rapide, des ajustements du métabolisme... Toutes ces adaptations ont un coût pour la plante en terme d’accumulation de biomasse. La recherche d’une tolérance à la sécheresse est donc celle d’un optimum entre des protections, qui ont un coût mais permettent à la plante de continuer à produire, et un objectif de production, qui est associé à une forte consommation d’eau au travers de grandes surfaces foliaires et l’ouverture des stomates, et accroit donc le risque de déshydratation des plantes lors des années sèches. Il serait donc plus juste de parler d’une optimisation que d’une tolérance à la sécheresse.
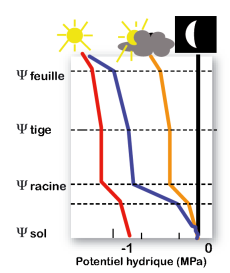
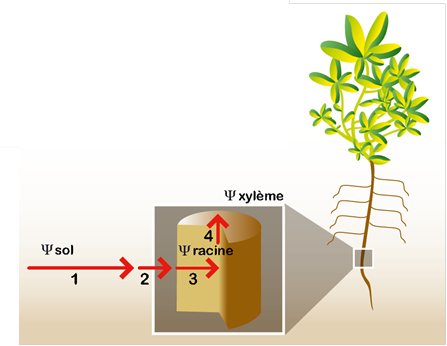
L’eau est d’autant plus transpirée par la feuille que la demande climatique est élevée (courbes en noir, orange et bleu). L’eau « coule » depuis le sol où elle est peu retenue (fort potentiel hydrique[1], ψsol) vers les feuilles où elle est davantage retenue (potentiel hydrique inférieur, ψfeuille). Chez une plante en déficit hydrique (courbe rouge), les stomates sont partiellement fermés. Ceci diminue le flux d’eau, et donc la différence de potentiel hydrique entre le sol et les racines. Les feuilles peuvent ainsi garder un potentiel hydrique acceptable bien que le sol soit sec. Cette fermeture stomatique est un processus adaptatif nécessaire à la survie de la plante. Elle a l’inconvénient de ralentir la photosynthèse et de causer l’échauffement de la feuille.
TOLÉRANCE OU PERFORMANCE
L’amélioration de la tolérance à la sécheresse est une voie étroite puisque la demande en transpiration est imposée par le climat, et que toute réduction de transpiration par réduction de surface foliaire ou par fermeture des stomates s’accompagne d’une réduction de la production de biomasse.
La recherche dans ce domaine a abordé la question avec plusieurs méthodes génétiques. La plus médiatisée est celle des plantes agronomiques génétiquement modifiées pour être tolérantes grâce à des transferts de gènes de plantes originaires de déserts. Ceci a effectivement permis d’améliorer la survie lors de sécheresses marquées, mais, à notre connaissance, jamais d’améliorer les rendements,. L’application pratique de ces recherches est l’obtention de plantes qui survivent, pour améliorer la pérennité d’une prairie ou éviter l’érosion, et non de plantes assurant une production importante en conditions sèches. Ce n’est que récemment que des organismes génétiquement modifiés (OGM) ont été développés pour améliorer les performances (et non la survie) de plantes soumises à des déficits hydriques. Elles sont menées par des compagnies semencières et des groupes publics, souvent en collaboration. Une variété d’OGM tolérante (maïs MON 87460) a été mise sur le marché après son autorisation aux États-Unis en décembre 2011. Son gain de production, d’environ 4-5 % par rapport à la variété d’origine en conditions sèches, est non négligeable mais très variable entre lieux et années. Il s’agit donc d’une amélioration, pas d’une transformation radicale permettant à la plante de produire en conditions de sécheresse intense.
La voie la plus utilisée jusqu’à présent par les firmes semencières est l’amélioration des performances des plantes en conditions habituelles de champ sans stress hydrique notable. Ceci a été efficace, avec un progrès génétique des rendements d'environ 1 % par an (Brisson et al., 2010, Campos et al., 2006). Les progrès en matière de rendement ont été similaires pour des plantes bien irriguées ou en déficit hydrique. Ceci a abouti à une amélioration spectaculaire des rendements du maïs en conditions sèches, avec un doublement entre 1950 et 2005, dans des essais comparant des variétés de différentes générations soumises à un même déficit hydrique (Cooper et al., 2009). L’analyse de ce progrès révèle qu’il ne concerne pas spécifiquement la réponse à la sécheresse. Il a été réalisé via des caractères simples, généralistes et relativement faciles à sélectionner. Il est probable que cette stratégie ne pourra pas être maintenue pendant une longue période car la marge de progrès se réduit.
La dernière voie consiste à rechercher des sources de tolérance à la sécheresse dans la diversité naturelle existant à l’intérieur de chaque espèce. Chacun des gènes d’une espèce a plusieurs modalités, appelées allèles, qui déterminent les caractéristiques des plantes. Il s’agit donc de rechercher, parmi la diversité existante, les allèles qui confèrent aux plantes des caractéristiques favorables en conditions de sécheresse. Ceci nécessite d’établir de grandes collections de plantes avec des origines diverses (issues de régions sèches en particulier), d’analyser la séquence du génome de chacune des plantes analysées et d’associer certains allèles avec des caractères d’intérêt. Ces allèles sont ensuite intégrés dans des variétés élites en vue d’une commercialisation.
QUELS CARACTÈRES POUR QUELS SCÉNARIOS DE SÉCHERESSE ?
La principale difficulté rencontrée dans ces approches est la variabilité du climat entre années et entre sites. Une plante bien adaptée à un scénario climatique ne l’est pas nécessairement à un autre. Un caractère, favorable dans un scénario, peut se révéler sans intérêt ou même nuisible dans un autre (Tardieu, 2011) (voir le tableau ci-dessous). Un réseau de 29 essais à travers l'Europe a récemment montré que ce qui s'applique aux caractères s'applique aussi aux gènes eux-mêmes. Les allèles de mêmes gènes peuvent affecter positivement ou négativement le rendement suivant les scénarios de sécheresse (Millet et al., 2016).
Intérêts et inconvénients de plusieurs caractères impliqués dans la tolérance à la sécheresse.
|
CARACTÈRES |
AVANTAGES |
INCONVÉNIENTS |
SCÉNARIO MAXIMISANT LES AVANTAGES |
SCÉNARIO MAXIMISANT LES INCONVÉNIENTS |
|
Une durée de vie plus courte ("esquive") |
Les plantes terminent leur vie avant que le sol ne soit trop sec |
La biomasse s’accumule pendant une durée |
Année très sèche |
Fin du cycle en conditions favorables |
|
Réduction de transpiration par fermeture stomatique ("évitement") |
La plante garde de l'eau pour la fin de la saison |
La plante produit moins de biomasse et de rendement |
Sécheresse en fin de saison |
Fin du cycle en conditions favorables |
|
Plus de racines |
Si le sol est profond, les plantes peuvent extraire plus d’eau |
Les racines coûtent cher en carbone, inutile si le sol est peu profond |
Sol profond |
Sol peu profond ou année humide |
|
Poursuivre la croissance en condition de sécheresse |
La plante produit plus de biomasse |
Risque de mort de la plante par manque d’eau |
Saison modérément sèche ou fin de saison humide |
Saison très sèche |
Une durée de cycle réduite est un caractère qui a été sélectionné depuis longtemps par l’évolution. Beaucoup d’espèces qui croissent dans le désert ont un cycle très court, qui leur permet de profiter d’une pluie pour produire rapidement des graines, lesquelles pourront attendre la prochaine pluie. Chez les plantes agronomiques, la réduction de durée du cycle (esquive) permet de terminer le cycle en conditions encore humides, même si l’année est sèche. Cependant, elle engendre une perte de biomasse en conditions relativement humides, puisque cette biomasse s’accumule pendant une période plus courte. Le choix de la durée du cycle est donc un pari sur le climat de l’année à venir, entre esquive (cycle court) et rendement potentiel maximum (cycle long). L’augmentation de la longueur ou de la biomasse du système racinaire peuvent apparaître comme des caractères favorables en conditions de sécheresse. C’est effectivement le cas en sol profond : les variétés qui ont un système racinaire profond captent davantage d’eau, ce qui leur permet d’accumuler plus de biomasse. Si le sol est peu profond en revanche, cette stratégie est néfaste parce qu’elle consiste à investir du carbone dans les racines sans que l’absorption d’eau soit améliorée (van Oosterom et al., 2016). Il est intéressant de noter que trois programmes d’amélioration génétique de plantes soumises à la sécheresse ont abouti, in fine, à diminuer la biomasse du système racinaire (Campos et al., 2006). Augmenter cette dernière n’a donc d’intérêt que dans certains milieux à sol profond.
Réduire la transpiration par voie génétique est possible en jouant sur la sensibilité des stomates au déficit hydrique. Il est notamment possible de manipuler la synthèse d’hormones (par exemple l'acide abscissique) qui servent de signalétique du stress entre les racines et les parties aériennes. Ceci permet d’économiser de l’eau (stratégie d’évitement), mais réduit la photosynthèse. Là encore, c’est une protection qui a un coût, et n’a d’intérêt que si le climat est particulièrement sec. À l’opposé, des allèles ont été identifiés par voie génétique, qui permettent aux feuilles de continuer à grandir même en cas de déficit hydrique prononcé (Welcker et al., 2011). Ils ne sont pas encore localisés précisément sur le génome, mais ils permettent de sauvegarder la capacité de photosynthèse de la plante. Ils peuvent aussi causer la mort de celle-ci par épuisement rapide de l’eau du sol si la sécheresse se prolonge. Chaque caractère de tolérance est ainsi associé à des scénarios précis de sécheresse. Dans une région donnée, le climat est imprévisible d’une année sur l’autre. Il faut au moins une dizaine d’années pour tester une plante et étudier ses caractères sous les climats les plus fréquents dans une région donnée, ce qui est bien sûr impossible. Pour remédier à ce problème, on peut recourir à des modèles de simulation qui permettent de prévoir le comportement d’un génotype dans n’importe quel scénario climatique, et donc de tester sur un grand nombre d’années si un génotype est plus adapté qu’un autre (Tardieu et Tuberosa, 2010).
[1] Le potentiel hydrique est une mesure de « l’avidité » en eau d’un compartiment du sol, de la plante ou de l’atmosphère. Voir Chap. I.2 et II.3
Sommaire :


