L’entrée de l’eau dans les racines
4.1 Rappels d’anatomie des racines
Une racine se présente généralement sous la forme d’un axe, plus ou moins cylindrique et ramifié, dont le diamètre décroît au fur et à mesure qu’on se rapproche de son extrémité. Le système racinaire assure trois fonctions principales : la nutrition hydrominérale, le stockage de réserves (souvent sous forme d’amidon) et enfin l’ancrage et le maintien mécanique de la plante. Ainsi, une racine comprend-elle des tissus de conduction, des parenchymes de réserve et des tissus de soutien.
Le système racinaire est un ensemble plus ou moins important de racines qui peuvent être très différentes du point de vue morphologique et anatomique. Chez les arbres par exemple, il existe plusieurs types de racines qui n’ont pas toutes le même rôle[1]. En simplifiant on peut les regrouper en deux grands types ; i) les racines d’ancrage et de réserve qui forment l’essentiel du système racinaire visible et qui peuvent s’étendre sur de grandes distances et à de grandes profondeurs ; ii) les racines fines, d’un diamètre compris entre un 0,1 et 1 mm, situées en périphérie, formant un chevelu à renouvellement plus ou moins rapide, souvent peu visible. Ce sont elles qui constituent la partie absorbante du système racinaire et dont il est question ici.
Les racines fines se trouvent à des profondeurs très différentes selon l’espèce, le type de sol, l’existence de compétition entre espèces, etc. Elles peuvent être confinées principalement dans la partie superficielle du sol (dans les 10 à 50 premiers cm) et y former un feutrage dense. Par exemple, Blaise et Garbaye, 1983 (cité dans Drénou, 2006), sur des hêtres, dans un sol brun acide (alocrisol) développé sur grès, ont dénombré 5 millions d’extrémités de racines fines par m2 dans les 30 premiers centimètres de sol, dont 94% dans les 10 premiers cm. Par contre dans une association peupliers – céréale d’hiver, le développement le plus important des racines fines se trouve à 180 cm de profondeur, sous les racines du blé[2].
Examinons la coupe d’une jeune racine dans le sens longitudinal (Fig. 39) puis dans le sens transversal (Fig. 40).
Longitudinalement on distingue les régions suivantes en partant de l’extrémité :
- la coiffe, région conique protégeant l’apex de la racine ;
- l’apex, zone par laquelle la racine grandit ;
- la région de développement des poils absorbants : elle est la région la plus perméable à l’eau, même si à ce niveau l’appareil conducteur n’est pas encore bien développé ;
- enfin une zone d’organogenèse dans laquelle des racines secondaires peuvent se développer.
Au dessus de la zone où se développent les poils absorbants, la couche de cellules la plus externe, appelé rhizoderme est plus ou moins subérisée c’est à dire recouverte de subérine hydrophobe.
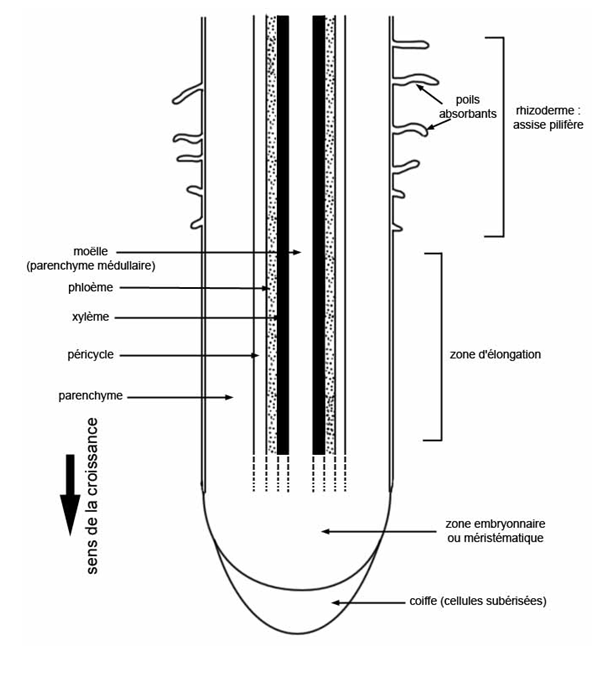
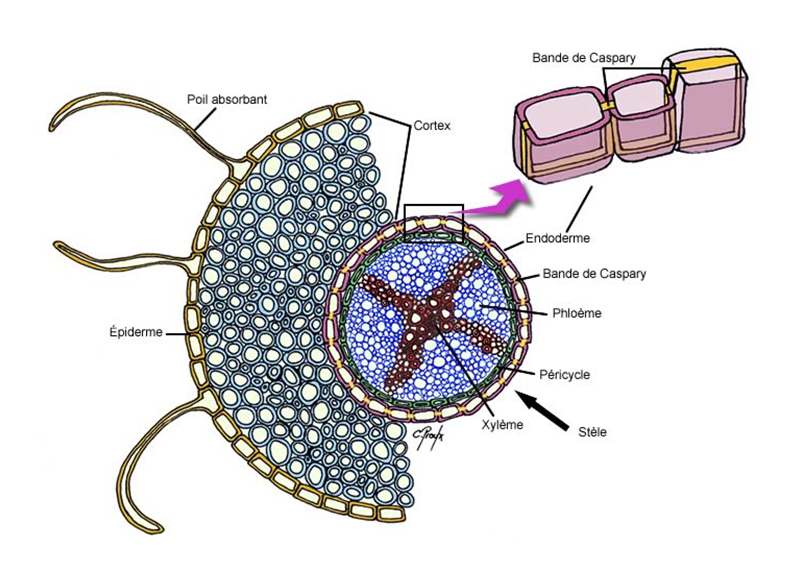
Figure 40. Coupe transversale d’une racine jeune. Source : https://www.cours-pharmacie.com/biologie-vegetale/architecture-vegetale.html
Transversalement (à partir de la zone d’élongation et au-dessus), de l’extérieur vers l’intérieur, une racine présente les couches suivantes (Fig. 40):
· une couche de revêtement externe, le rhizoderme (appelé aussi assise pilifère), assez analogue à l’épiderme des parties aériennes (mais sans stomates ni cuticule), perméable à l’eau. Certaines de ces cellules sont transformées en poils absorbants, sortes de filaments unicellulaires de quelques millimètres de longueur et de quelques centièmes de mm de diamètre. Ils peuvent être recouverts d’un mucilage rétenteur d’eau. Ils se situent dans une zone très précise des racines, zone juste au-dessus de la coiffe, partie en croissance de la racine.
· juste au-dessous du rhizoderme un ensemble de plusieurs couches cellulaires formant le parenchyme cortical (ou cortex) plus ou moins large pouvant servir de zone de stockage de réserves. Elle est souvent la zone la plus développée de la racine.
· juste au-dessous du rhizoderme, une assise cellulaire dont les parois sont ceinturées par une couche de subérine d’abord, puis de cellulose. Ce revêtement forme une barrière au passage de l’eau. Il est appelé la bande de Caspary.
· enfin, la partie la plus centrale de la racine, appelée pour cette raison « cylindre central ». Cette région commence par une couche de cellules appelée péricycle, adossée à l’endoderme (côté centre de la racine). Le péricycle est la première couche cellulaire du cylindre central. Celui-ci contient les éléments conducteurs de la sève brute (le xylème) et de la sève élaborée (le phloème).
4.2 Le passage de l’eau, du sol dans les racines ; la bande de Caspary
Dans la majorité des cas une racine ne peut fonctionner, et en particulier absorber l’eau du sol, que si au moins une partie de sa surface se trouve dans une atmosphère humide, voire dans une eau contenant assez d’oxygène. Cela signifie qu’en conditions normales non pas toute, mais seulement une partie plus ou moins importante de la surface de la racine se trouve en contact avec l’eau liquide d’un pore du sol. Seules certaines zones de la surface de la racine sont susceptibles d’absorber l’eau du sol ; la région la plus perméable est celle où se trouvent les poils absorbants, mais elle n’est pas la seule laissant entrer l’eau.
Dans un grand nombre de cas, la partie la plus distale des racines est entourée par une couche très particulière de sol qu’on appelle la rhizosphère, dans laquelle se situe en particulier le mycélium des mycorhizes[3]. Cette couche de sol aussi est très modifiée par l’activité racinaire (re-largage de dioxyde de carbone, sécrétions de sucres, d’acides aminés, substances diverses…).
En gardant en tête le fait que la zone de contact sol-racine est très hétérogène et variable, les étapes du passage de l’eau de l’entrée dans la racine jusqu’au cylindre central sont les suivantes :
1- Le trajet du sol au rhizoderme. On entend encore dire parfois que l’eau du sol pénètre dans une racine par les seules cellules de la couche cellulaire la plus extérieure, le rhizoderme, transformées en poils absorbants, mais ce n’est pas exact. La présence de poils absorbants[4] est limitée à une zone assez courte de la racine, située juste au dessus de la zone d’élongation. Cette zone de poils absorbants grandit ou se déplace avec la croissance de la racine. La durée de vie des poils absorbants est très variable : de quelques jours à quelques semaines, parfois plus. On les rencontre chez un très grand nombre d’espèces mais ils sont absents chez d’autres (pacanier, arachide, nombre de conifères, les plantes aquatiques en général). Leur densité est très variable, selon les espèces et les conditions du sol (de quelques centaines à quelques milliers par cm2,[5]). Leur développement est favorisé par une bonne aération et une humidité du sol élevée[6], proche de la capacité au champ[7] (voir Chap. I.4). On leur a longtemps attribué un rôle important, voire prépondérant dans l’absorption racinaire. Les mesure montrent que ce n’est pas du tout certain[8] [9] et d'ailleurs nous avons indiqué que certaines espèces n'en possédaient pas. Mais il est vrai que leur présence augmente considérablement la surface d’absorption des racines ; mais il n’est pas certain que la quantité d’eau absorbée par une racine ou un système racinaire soit toujours proportionnelle à leur surface.
Des observations montrent que nombre d’espèces, ligneuses en particulier (des pins, le troène, le houx ou le citronnier), survivent en conditions de sols secs ou froids alors que leurs racines ne présentent que des surfaces épidermiques non subérisées réduites, que leur croissance en longueur a cessé et que la transpiration peut pourtant rester élevée. Si ces espèces ne meurent pas c’est qu'il y a absorption d'eau [10] [11] [12] dans les zones subérisées des racine matures. Meidner et Sheriff (1976) suggèrent même qu’en cas de stress hydrique, ces zones subérisées jouent un rôle plus important dans l’alimentation hydrique que les zones non subérisées.
2- La traversée du cortex. Sur ce trajet, les deux voies apoplastique et symplastique peuvent être utilisées. La voie apoplastique est ouverte partout, sauf aux endroits où se trouve la bande de Caspary. Celle-ci se présente sous la forme d’un dépôt de lignine et ou de subérine localisé dans les parois radiales et parfois transversales des cellules. Du fait que lignine et subérine sont des composés hydrophobes on a longtemps crû que cette structure était imperméable à l’eau qui, de ce fait était obligée de passer par l’intérieur des cellules.
Certes, la bande de Caspary est présente dans les deux premières couches de cellules (rhizoderme et l’exoderme) ainsi que dans l’endoderme. Cependant son efficacité en tant que barrière à l’eau n’est pas systématique : elle n’est pas imperméable dans toutes les cellules de ces deux couches cellulaires [13]. Il faut aussi tenir compte du fait que la bande de Caspary évolue dans le temps comme son degré de barrage pour l’eau et qu’elle est interrompue au niveau où se développent les racines secondaires. Aujourd’hui, on peut donc tout juste dire que les dépôts de subérine dans les parois offrent certainement un barrage total dans certains cas ; mais le plus souvent ce barrage ne fait que ralentir, plus ou moins, le passage de l’eau.
La seconde voie utilisable, la voie symplasmique, est constituée par le continuum des cytoplasmes interconnectés par les plasmodesmes.
3) L’entrée dans le cylindre central. Les cellules de celui-ci, souvent plus petites que celles du cortex, forment une enveloppe serrée autour des éléments conducteurs qui convoient l’eau vers la partie aérienne. Là encore les modalités précises de ce passage sont multiples et dépendantes notamment des formes de développement et de maturité des éléments conducteurs. Lorsque la sève brute arrive dans l’appareil conducteur, son trajet jusqu’alors globalement horizontal, devient vertical, vers la partie aérienne.


