Importance des relations hydriques pour l’expansion cellulaire
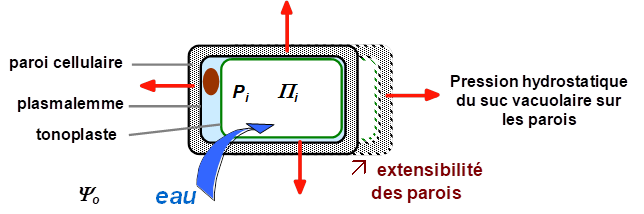
Alors que chez les animaux la croissance se fait via la prolifération des cellules, chez les végétaux, la croissance se fait grâce à deux processus, la prolifération des cellules et surtout leur expansion volumique (l’auxèse). Cette expansion cellulaire ou grandissement cellulaire consiste en une absorption massive d'eau couplée à une extension des parois pectocellulosiques. Par ce processus, schématisé ci-dessous, le volume cellulaire est augmenté de 10 à 100 fois, selon les types cellulaires et les espèces.
Le modèle de Lockhart
En 1965 Lockhart a établi un modèle qui décrit les différents paramètres biophysiques contrôlant le grandissement d’une cellule. Il a établi la première partie de son modèle sur cette observation : la cellule végétale se comporte comme un osmomètre (presque) parfait. Le flux d’eau entrant est passif, induit par la différence de potentiel hydrique entre l'intérieur de la cellule et le milieu extérieur. La variation relative du volume cellulaire, correspondant à ce flux d'eau entrant dans la cellule en croissance, peut être ainsi exprimée :
![]() (éq. 14)
(éq. 14)
avec : ![]()
où L est la conductance hydraulique volumique de la cellule, qui dépend de V le volume cellulaire, A sa surface, et Lp la conductivité hydraulique de la membrane plasmique. Le coefficient de réflectivité membranaire s peut raisonnablement être considéré égal à 1 (membrane hémiperméable). De plus, si on considère que la pression de l’environnement extérieur Po est nulle (pas de surpression ou dépression par rapport à la pression atmosphérique :
![]() (éq. 15)
(éq. 15)
Le grandissement cellulaire dépend donc de l'existence d'une différence de potentiel hydrique entre la cellule en croissance et le milieu environnant. Si celle-ci s'annule, l'entrée d'eau s'arrête et la croissance cesse. La conductivité hydraulique membranaire peut également réguler l'entrée d'eau dans la cellule et son rôle sera discuté ci-après. Le flux d'eau entrant dans la cellule en croissance constitue l'aspect hydraulique du grandissement cellulaire.
La deuxième partie du modèle de Lockhart considère l’aspect mécanique : pour que puisse augmenter significativement le volume cellulaire, il faut une extension irréversible des parois (Il ne s’agit plus d’extension réversible due à de l’élasticité des parois, comme dans le diagramme de Höfler). D’autre part, la vitesse d’accroissement du volume est proportionnelle à la pression de turgescence. Ainsi :
![]() (éq. 16)
(éq. 16)
où f et Y représentent l’extensibilité des parois, incluant un coefficient de proportionnalité et un seuil minimum de pression. Cette relation est bien visible sur l’exemple de la Fig. 8 (issue de Pritchard, 1994).
Le modèle de Lockhart intègre les aspects hydraulique et mécanique du grandissement cellulaire. En combinant les équations 15 et 16, on obtient :
![]()
Ainsi, selon le modèle de Lockhart, l’augmentation relative du volume cellulaire V dépend :
- du gradient de potentiel osmotique entre l’intérieur et l’extérieur de la cellule DPi,o,
- de l’extensibilité des parois (f et Y),
- de la conductance hydraulique volumique cellulaire L
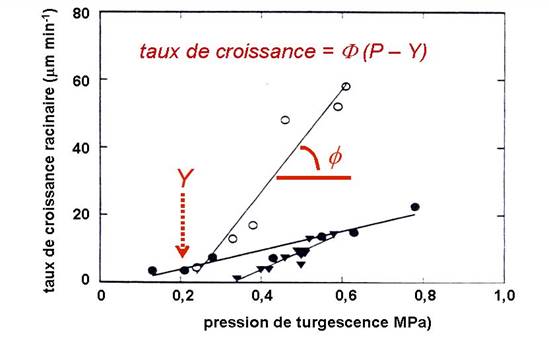
Figure 8. Taux de croissance racinaire relatif de différentes espèces en fonction de la pression de turgescence ; des racines de maïs (o) et de blé (·) ont été plongées dans une série de solutions de mannitol. Des racines de pois (▼) ont poussé dans des sols de différentes densités (Pritchard 1994).
Potentiel hydrique induit par la croissance cellulaire
La pression de turgescence est le moteur de l’expansion, en générant de la tension dans les parois qui va permettre leur extension. L’extensibilité des parois et la conductance hydraulique membranaire en sont deux rhéostats.
L’extensibilité des parois quantifie le relâchement pariétal. Ce relâchement a été perçu et analysé comme un comportement viscoélastique des parois, même si l’idée de processus chimiques, impliquant que la vitesse d’expansion cellulaire dépend de vitesses de réactions biochimiques, a été émise très tôt par Lockhart. Ce relâchement des parois est à l’origine de la baisse de la pression de turgescence et donc du gradient de potentiel hydrique nécessaire au flux d’eau entrant dans la cellule en croissance (Cosgrove, 1993).
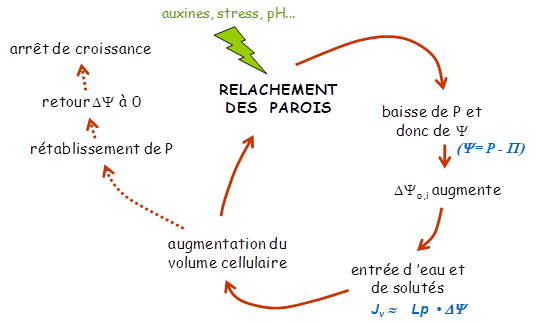
La limitation de la croissance par la conductance hydraulique a fait régulièrement débat. L’intensité du gradient de potentiel hydrique entre la source (les cellules matures, l’apoplasme environnant) et le puits (les cellules en croissance) dépend de la conductance hydraulique de la membrane / du tissu entre la source d’eau et les cellules en croissance. Elle reflète donc l’importance de cette limitation hydraulique. Le potentiel hydrique mesuré dans la zone de croissance de l’hypocotyle de ricin et celle de la feuille de fétuque élevée a été trouvé de 0.15 à 0.30 MPa plus négatif que celui des zones matures ou du xylème environnant, indiquant une réelle limitation hydraulique de l’expansion cellulaire. Au contraire, dans les apex racinaires, aucun gradient de potentiel n’a été détecté entre cellules en croissance et cellules matures. Cependant, ce dernier résultat est peut-être un artefact. Les manipulations nécessaires pour fixer la racine dans la cuve (pour la mesure directe de P et P cellulaire) provoquent très certainement un arrêt de croissance et la turgescence et le potentiel osmotique ont probablement été mesurés dans des cellules dont la croissance était stoppée. Une limitation hydraulique de l’expansion dans l’apex racinaire n’est donc pas exclue. Plus récemment, l’origine de l’eau alimentant les cellules en expansion dans l’apex de racine en croissance et les conséquences sur les gradients de potentiel ont été revues. La part de la source phloémienne semble importante et les gradients de potentiel hydrique significatifs, soutenant l’hypothèse d’une possible limitation hydraulique de la croissance dans cet organe. Parallèlement, la découvertes des aquaporines, des protéines intra-membranaires facilitant les flux d’eau à travers les membranes (pores), a induit une re-visitation des processus physiologiques impliquant des flux d’eau, y compris l’expansion cellulaire. Plusieurs études ont mis en évidence l’implication d’aquaporines dans le contrôle des processus de croissance dans les zones de croissance de feuilles et de racines, apportant un élément de plus en faveur d’une limitation hydraulique de l’expansion et surtout l’idée d’une potentielle régulation de la croissance via la quantité et l’activité de ces pores transmembranaires. La multiplicité de ces protéines (famille multigénique) et leur potentielle redondance rendent complexe la détermination de leurs implications respectives.
L’expansion cellulaire a ainsi été très souvent analysée comme potentiellement limitée par l’extensibilité des parois et/ou par le flux entrant d’eau. D’autres études ont mis en évidence que le flux de solutés, nécessaire au maintien de la pression osmotique, doit être considéré comme un autre facteur potentiellement limitant. De plus, dans le contexte d’organismes pluricellulaires complexes, il serait nécessaire de replacer l’expansion cellulaire au sein de son tissu. La pression de turgescence, le flux d’eau entrant et la vitesse de croissance de la cellule en expansion dépendent de la pression dans les parois et le xylème avoisinant (apoplasme), cette dernière dépendant de l’état de transpiration de la plante.éines. Une mole de protéine de 1kDa pèse 1000 g.
Sommaire :
Dans le chapitre :
- Introduction : la cellule végétale vivan...
- Le potentiel hydrique de la cellule et s...
- Relation potentiel hydrique – volume cel...
- L’osmorégulation
- Les transferts d’eau transmembranaires
- Importance des relations hydriques pour ...
- Mesure des composantes du potentiel hydr...
- Références bibliographiques


