La double circulation des sèves
Au niveau de la plante entière, les échanges gazeux foliaires (photosynthèse et transpiration) sont à l'origine des deux flux de sève qui circulent en sens opposé dans des structures distinctes :
- Flux de sève brute : circule dans les vaisseaux du bois (xylème) dont la structure étudiée au chapitre II.2 permet le passage des débits très importants d’eau imposés par la demande transpiratoire. Son trajet débute par l’absorption de l’eau du sol par les racines fines. D’un point de vue chimique c’est une solution minérale très peu concentrée, dont la composition reste proche de celle de l'eau du sol.
- Flux de sève élaborée : extrêmement riche au contraire en éléments dissous, sucres notamment, d’où une viscosité importante. Elle circule globalement vers le bas et très lentement dans le phloème, tissu spécialisé composé de vaisseaux très fins (tubes criblés). Sa fonction est de répartir dans l’ensemble de la plante les photoassimilats issus des feuilles, ainsi que de nombreuses autres molécules dissoutes à moindre concentration.
D'un point de vue anatomique, xylème et phloème sont souvent proches tout en restant parfaitement distincts (figure 4 et chapitre II.2). Ils ont par ailleurs des caractéristiques fonctionnelles contrastées adaptées aux propriétés des deux sèves qu'ils véhiculent. Ainsi, le xylème fonctionnel (bois), est un ensemble de vaisseaux de diamètre assez fort qui offrent au total une section importante pour la circulation de la sève brute. Il n'est constitué que de tissus inertes dont la mise en place s'effectue progressivement par un processus de « maturation » au cours duquel les cellules initiales meurent par suite d'un dépôt de lignine imperméable (paroi secondaire). Au contraire, le phloème correspond à un ensemble de cellules vivantes hyperspécialisées, les tubes criblés. En raison d’une extrême réduction de leur équipement intracellulaire (absence de noyau et de vacuole, à peine quelques organites résiduels, cytoplasme confondu avec la solution de sève élaborée circulant d’une cellule à la suivante, leur métabolisme est assuré par les cellules compagnes qui leur sont étroitement associées. Ils ont cependant conservé une caractéristique essentielle des cellules vivantes : la membrane plasmique qui les délimite et qui joue un rôle capital dans la fonction de conduction (Van Bel, 2003).
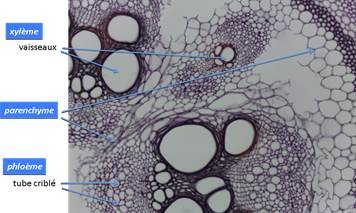
Figure 4. Microphotographie d’une coupe transversale de tige de vigne. On reconnait les vaisseaux du bois (xylème), structures mortes où circule la sève brute et un certain nombre de structures vivantes : tubes criblés du phloème (dans lesquels circule la sève élaborée), et les parenchymes, siège de nombreuses fonctions physiologiques (Source : BCPST lycée Fénelon, http://www.bcpst.eu/spip.php?article127).
Rappelons la distinction fondamentale entre apoplasme et symplasme chez les végétaux (figure 5):
- l'apoplasme désigne tout le compartiment cellulaire situé en position externe par rapport à la membrane cytoplasmique. L'eau apoplasmique est retenue par le phénomène purement physique de capillarité dans les structures microporeuses des parois cellulosiques (micro-fibrilles) et dans la macroporosité des méats intercellulaires. Sauf exception, l’eau apoplasmique est très peu chargée en solutés (pression osmotique négligeable) et son potentiel hydrique (Y) est négatif (cf. chapitre I.2). L’ensemble du xylème est rattaché à l’apoplasme, la sève brute est ainsi une eau apoplasmique.
- le symplasme désigne le compartiment cellulaire situé en position interne par rapport à la membrane cytoplasmique et à la paroi. Au symplasme sont rattachés les vacuoles et tous les organites qui sont le siège des fonctions physiologiques et biochimiques de la cellule. D'une façon générale l'eau symplasmique, très chargée en solutés (pression osmotique élevée) est soumise à une forte pression hydrostatique (pression de turgescence) par suite de la tendance à l'entrée de l'eau pure par osmose (cf. chapitre I.2). C'est la paroi cellulosique qui par son élasticité permet à la cellule de résister à des pressions de turgescence souvent très élevées (> 10 bars ou 1 MPa). De ce point de vue la cellule se comporte comme un osmomètre. L'état hydrique du symplasme est décrit par deux variables : la pression osmotique (P) et la pression hydrostatique ou de turgescence (P). L’ensemble du phloème est rattaché au symplasme, la sève élaborée est une eau symplasmique.
Les deux compartiments sont séparés par la membrane plasmique que l’on peut assimiler à une membrane semi-perméable, c’est à dire qu’elle est traversée plus ou moins librement par les molécules d’eau alors que les solutés ne la traversent que par le biais d’un contrôle physiologique. A l’échelle cellulaire on peut admettre que le symplasme et l'apoplasme sont le plus souvent proches de l’équilibre hydrique :
![]() (équat. 5)
(équat. 5)
Mais cette condition n’est pas obligatoire : en l’absence d’équilibre, un flux d’eau doit traverser la membrane cytoplasmique dans un sens ou dans l’autre. C’est ce qui se passe en particulier de part et d’autre de la membrane au niveau des tubes criblés et des cellules compagnes, où du fait d’un chargement/déchargement continuel du phloème, l’équilibre osmotique n’est jamais atteint.
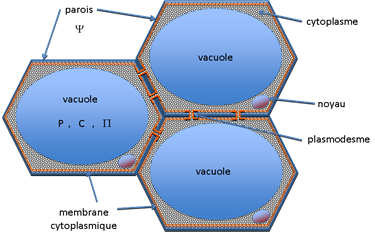
Figure 5. Apoplasme et symplasme des cellules végétales : deux compartiments hydriques disjoints et intriqués, séparés par la membrane plasmique. L’apoplasme regroupe les parois cellulosiques et les espaces poreux intercellulaires qui sont des structures inertes. L’eau apoplasmique est généralement sous tension et le potentiel hydrique Y négatif. La membrane cytoplasmique (représentée en orange sur la figure) est semi-perméable de sorte que la cellule se comporte comme un petit osmomètre. Le symplasme regroupe l’ensemble des structures vivantes situées à l’intérieur de la membrane cytoplasmique (vacuole, cytoplasme, noyau et autres organites). Les symplasmes de cellules adjacentes sont en continuité par l’intermédiaire des plasmodesmes.
II.1 LES MECANISMES DE CIRCULATION DES DEUX SEVES
Sève brute et sève élaborée sont en continuité avec les compartiments d’eau apoplasmique et symplasmique respectivement (cf. chapitre II.1), présents dans toutes les cellules végétales vivantes. Ainsi la sève brute passe librement des vaisseaux terminaux (extrémité des nervures) dans les parois pecto-cellulosiques des parenchymes foliaires notamment. De même, les éléments dissous de la sève élaborée véhiculés par le phloème circulent par la voie symplasmique (via les plasmodesmes, figure 5) pour alimenter les moindres puits de carbone cellulaires.
Le flux de sève brute
Le flux de sève brute compense la perte d’eau occasionnée par la transpiration foliaire. Lorsque cette compensation n’est pas totale, les réserves d’eau présentes dans les feuilles elles-mêmes ou dans les tissus sous-jacents sont mises à contribution. Lorsque le dessèchement des tissus devient trop important, la régulation stomatique intervient pour limiter les pertes en eau ce qui a pour conséquence une réduction simultanée de la photosynthèse.
Ascension de la sève brute
On s'est longtemps interrogé sur l'ascension de la sève dans les plantes, particulièrement dans les grands arbres. La sève est-elle poussée activement au niveau des racines (poussée racinaire par exemple) ou est-elle tirée par le haut ? Ces deux options ne pourraient-elles pas coexister ? Il est clair aujourd'hui que la poussée racinaire est un phénomène d'intensité beaucoup trop faible pour être impliqué de façon significative dans les flux d'eau qui transitent à travers les plantes du fait de la transpiration pendant la journée.
Le consensus est bien réalisé actuellement en faveur de la théorie de la « tension-cohésion », malgré certaines lacunes dans notre connaissance/compréhension des mécanismes qu’elle implique (chapitre II.6). Dans cette théorie, la transpiration est le moteur de l'ascension de la sève brute. Tout se passe comme si la sève était « aspirée » ou tirée au niveau des feuilles, cette aspiration se transmettant de proche en proche jusqu'aux racines grâce à la cohésion de l'eau.
Le flux ascendant de la sève brute s'accompagne d'une importante perte de charge dans les vaisseaux du fait de la viscosité de l'eau (au même titre que dans n'importe quelle canalisation), perte de charge qui s‘ajoute à la diminution de pression hydrostatique liée à l’altitude (environ 104 Pa m-1) et qui se traduit par une diminution de potentiel hydrique lorsqu’on parcourt la plante depuis la base jusqu’au sommet (YB < YA sur la figure 6). L'eau du sol étant le plus souvent déjà à une pression négative (tension) la sève se trouve donc à des pressions encore plus basses (= plus négatives) et ceci d'autant plus que l'on se rapproche des feuilles du sommet de la plante et que le flux de transpiration est intense. C'est le volet « tension » de la théorie, volet abondamment confirmé par les mesures de potentiel hydrique effectuées sur des plantes en conditions de transpiration (chapitre II.6).
Le volet « cohésion » s'il rend bien compte de la transmission de proche en proche du mouvement de sève, pose encore problème. Il est difficile en effet d'expliquer comment la colonne de sève mise en mouvement dans les vaisseaux du xylème, soumise comme on vient de le dire à des tensions extrêmement fortes, résiste à la cavitation. Pourquoi et comment n'est-elle pas systématiquement interrompue par des bulles de gaz (air, vapeur d’eau), ce qui abolirait l'intégrité des vaisseaux xylémiens et leur aptitude à conduire la sève ? Cette question est encore insuffisamment comprise. Lire à ce sujet le complément C3 « cohésion de l’eau » en fin de chapitre.
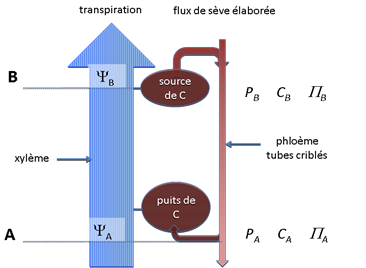
Figure 6. Schéma d’organisation d’une plante (hypothèse de Münch). On a figuré le flux de sève brute déterminé par la transpiration qui s’accompagne d’un gradient de potentiel hydrique total dans le xylème décroissant vers le haut de la plante (YA>YB). L’activité photosynthétique d’une feuille source (en B) conduit au chargement de sucres solubles dans le phloème alors que le puits en A en provoque le déchargement. Il en résulte un gradient de pression de turgescence le long du phloème (PB > PA), responsable du flux basipète de sève élaborée (mass flow).
Le flux de sève élaborée
En l'absence de « pompes » physiologiques actives qui seraient susceptibles de mettre en mouvement la sève élaborée et d'en assurer la distribution dans tous les tissus, quel mécanisme permet cette redistribution ?
Au niveau des organes sources (ex. feuilles photosynthétiques) le « chargement du phloème » c'est à dire le déversement des sucres solubles dans les tubes criblés, mécanisme actif, conduit à des concentrations de sucres élevées dans ces organes sources alors que des concentrations beaucoup plus basses se développent dans les zones de déchargement du phloème en réponse à l'activité des puits (CB >CA sur la figure 6). Ce gradient de concentration le long des tubes criblés entre zones source et puits ne suffirait pas à entrainer un flux important de sucre par simple diffusion. C'est un mécanisme beaucoup plus puissant qui est à l'œuvre : un transport par convection (mass flow) envisagé par Münch (1930). La condition de l’équilibre osmotique (Y = P - P, cf. chapitre I.2) impose en effet que la pression de turgescence dans les tubes criblés soit grosso modo proportionnelle à la concentration en sucres. Ainsi le gradient de concentration qui s'installe dans le phloème par suite des activités source/puits s'accompagne-t-il d'un gradient équivalent de pression. C'est ce gradient de pression (PB > PA, figure 6) qui provoque la mise en mouvement de la sève élaborée vers la base de la plante, entrainant celle des sucres des sources vers les puits. Cette explication est unanimement admise aujourd’hui.
Le flux de sève élaborée apparaît alors comme un phénomène purement passif. C’est l’ensemble des activités des sources et des puits – elles mêmes contrôlées par des régulations hormonales ou autres –, qui déterminerait à lui seul l’orientation des flux de sucre, en fonction des besoins des différents organes et des résistances hydrauliques.
II.2 Lois de circulation de la sève : formulation ohmique, pertes de charge, résistances
La figure 6 présente un schéma simplifié de la circulation des sèves, schéma que nous serons amenés à analyser et étudier de façon détaillée dans ce qui suit. La question se pose tout d’abord de savoir dans quelle mesure les lois ordinaires de la mécanique des fluides peuvent s’appliquer à la circulation de la sève dans les végétaux.
Si nous prenons l’exemple du xylème secondaire des angiospermes, tissu conducteur qui assure l’essentiel du flux de la sève brute, il est constitué d’un ensemble de très nombreux vaisseaux ou trachéides du bois dont le rayon est généralement compris entre la dizaine et la centaine de microns. Ces éléments ont pu être assimilés à un ensemble de tubes capillaires.
On sait que le débit Q qui circule en réponse à une différence de pression DP entre l’entrée et la sortie d’un capillaire de rayon r et de longueur l est donné par la loi de Poiseuille :
![]() (équat. 6)
(équat. 6)
où m est la viscosité dynamique du fluide
Cette loi montre principalement que le débit est :
- proportionnel à la perte de charge DP
- proportionnel à r4, c’est à dire au carré de la section de passage.
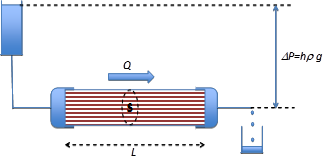
Figure 7. Dispositif expérimental permettant de mesurer la résistance hydraulique d’un rameau. Connaissant le débit Q et la perte de charge DP=hrg. La résistance est donnée par R= DP/Q
Etudions maintenant le flux d’eau qui traverse un rameau à l’aide du dispositif schématisé figure 7, en faisant varier la différence de pression DP=hrg entre les deux extrémités, puis comparons des rameaux de longueur l différente. L’expérience montre que le débit d’eau Q est proportionnel à la différence de pression imposée DP et inversement proportionnel à la longueur l du rameau. Le rameau se comporte du point de vue hydraulique comme un ensemble de tubes capillaires parallèles, obéissant à la loi de Poiseuille. Cette expérience permet notamment de définir la résistance hydraulique du rameau R :
![]() (Pa m-3 s) (équat. 7)
(Pa m-3 s) (équat. 7)
Cette définition est formellement analogue à celle de la résistance électrique de la loi d’Ohm (R=DV/I) où DV est la différence de potentiel électrique aux bornes de la résistance et I l’intensité du courant. C’est ce que l’on appelle couramment l’analogie ohmique. L’analyse physique faite à partir de la loi de Poiseuille met en évidence les deux principaux facteurs qui déterminent la résistance hydraulique du xylème :
· la section de passage effective (somme des sections de passage offertes par les vaisseaux du bois)
· mais surtout la distribution des diamètres des pores (appelés « lumières ») puisque nous avons vu que le débit qui traverse un tube capillaire est proportionnel à la puissance quatrième du rayon des pores.
Transferts d’eau liquide dans les systèmes apoplasmique et symplasmique
Les notations utilisées correspondent à celles de la figure 6.
Xylème et apoplasme : rôle du potentiel hydrique
La grande majorité des travaux sur la circulation de l'eau dans la plante concerne le flux transpiratoire globalement vertical qui part du sol et des racines et se termine au niveau des feuilles. Van den Honert (1947) et de multiples auteurs par la suite ont pu croiser sur le terrain des mesures de flux de sève parallèlement avec des mesures de potentiel hydrique. Ils ont démontré que :
1. le flux de sève brute dans une plante qui transpire est associé à une chute de potentiel hydrique Y entre le sol et les feuilles du sommet,
2. cette chute de potentiel est proportionnelle à l’intensité du flux de sève.
La sève brute apparaît ainsi comme un fluide visqueux « normal » dont la circulation dans les éléments poreux du xylème s’accompagne de « pertes de charge », comme dans un système hydraulique ordinaire. Cette observation et les nombreuses expérimentations menées tant en conditions de laboratoire que sur le terrain ont validé l’usage de la notion de résistance hydraulique pour caractériser les propriétés du tissu conducteur. Le formalisme ohmique décrit la relation entre flux de sève et chute de potentiel hydrique :
![]() (équat. 8)
(équat. 8)
avec : ![]() = flux volumique d’eau (m3 s-1)
= flux volumique d’eau (m3 s-1)
YA , YB = les potentiels hydriques aux points A et B respectivement (Pa).
La relation 8 montre que l’on peut mesurer![]() si l’on dispose simultanément d’une mesure du flux de sève et des valeurs du potentiel hydrique aux extrémités A et B.
si l’on dispose simultanément d’une mesure du flux de sève et des valeurs du potentiel hydrique aux extrémités A et B.
Phloème et symplasme : rôle de la pression de turgescence
Les sucres et autres substances dissoutes sont majoritairement entrainés par le flux de sève élaborée et circulent dans les tubes criblés du phloème. Ce mouvement se fait dans le sens opposé à celui de la sève brute c’est à dire à contre gradient de potentiel hydrique Y. Il est donc évident que le Y n’intervient pas directement dans le flux de sève élaborée. L’hypothèse de Münch (1930) déjà évoquée et largement admise, explique le flux de sève élaborée comme conséquence du gradient de pression qui s’établit dans les tubes criblés par suite des phénomènes de chargement/déchargement de sucres dans le phloème résultant des activités sources et puits des organes (Thompson & Holbrook, 2003).
Il est logique d’appliquer à la sève élaborée le même principe qu’à la sève brute (formulation ohmique) en remplaçant le potentiel hydrique Y par la pression de turgescence P. Le flux de sève élaborée qui parcourt un tronçon élémentaire AB s’écrira donc :
![]() (équat. 9)
(équat. 9)
PA et PB sont les pressions de turgescence en A et B respectivement. La résistance hydraulique ![]() qui apparait ici est celle des tubes criblés. D’un point de vue expérimental, cette résistance n’est pas mesurable facilement. Des valeurs approchées peuvent néanmoins être obtenues en modélisant le flux visqueux de la sève dans les tubes criblés par application de la loi de Poiseuille, ou plus indirectement en combinant des informations sur la vitesse de la sève, les flux de sucres ainsi que le potentiel hydrique.
qui apparait ici est celle des tubes criblés. D’un point de vue expérimental, cette résistance n’est pas mesurable facilement. Des valeurs approchées peuvent néanmoins être obtenues en modélisant le flux visqueux de la sève dans les tubes criblés par application de la loi de Poiseuille, ou plus indirectement en combinant des informations sur la vitesse de la sève, les flux de sucres ainsi que le potentiel hydrique.
Transfert d’eau entre apoplasme et symplasme
On peut supposer que symplasme et apoplasme sont généralement proches de l’équilibre hydrique, mais il peut arriver que le potentiel hydrique de l’apoplasme Yapoplasme soit sensiblement différent de celui du symplasme (Ysymplasme = P-P). Dans ce cas un transfert d’eau s’établit entre les deux compartiments à travers une résistance hydraulique particulière qui traduit la perméabilité à l’eau de la membrane cytoplasmique.


