Nécessité de quantifier séparément Evaporation du sol et Transpiration des plantes
L’estimation séparée de l’évaporation directe du sol (E) et de la transpiration des plantes (T) est un prérequis pour l’étude de l’adaptation des systèmes de culture au manque d’eau. Les méthodes pour effectuer cette séparation sont assez diverses : modélisation des processus (Sanchez et al. 2015 ; Fandino et al. 2015), utilisation de micro-lysimètres (Villalobos & Fereres, 1990) ou de la discrimination isotopique (Wang et al. 2015 ; Yidana et al. 2016), mais elles sont assez difficiles à mettre en œuvre en routine. Plus simplement, Hanks et al. (1969) ont proposé, à partir de séries de données reliant la biomasse des cultures à la récolte (B) à la quantité d’eau consommée (E+T), que l’abscisse à l’origine de la régression B=f(E+T) pouvait être une estimation de E. Kunrath et al. (2018) ont adapté cette approche statistique à l’étude dynamique de la consommation en eau d’une culture au cours de son cycle (Figure 1).
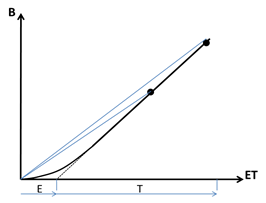
Figure 1 : Représentation de la dynamique d’accumulation de biomasse d’une culture en fonction de la quantité d’eau consommée (évapotranspiration ET) d’après Kunrath et al. 2018. L’extrapolation de la partie linéaire de la courbe B = f(ET) permet d’évaluer les deux composantes de ET : transpiration (T) et évaporation (E). Le rapport B/ET représentant l’efficience totale de l’eau consommée EffET (pente des droites) est donc dépendant des niveaux de biomasse (B) et de consommation en eau (ET) atteints par la culture à la récolte.
La régression linéaire entre B et ET calculée lorsque l’indice foliaire de la culture est suffisamment élevé pour que E devienne négligeable devant T peut alors être extrapolée et permet d’estimer la part d’évaporation du sol (E). Le réalisme de cette estimation dépend de deux conditions: (i) que la composante E devienne réellement négligeable lorsque l’interception du rayonnement incident devient proche de son maximum ; et (ii) que la pente de la régression dB/dET soit bien constante tout au long de la période de culture. En effet, une variation de cette pente en cours de culture affecterait fortement la détermination de l’abscisse à l’origine et donc l’estimation de E. Pour le premier point, il est évident que l’on ne peut jamais affirmer que E=0 car la composante non radiative de E sera toujours active du fait du déficit de vapeur d’eau de l’atmosphère. Il y a donc un risque de sous-estimation de E notamment avec des sols maintenus humides en surface (régime irrigué) et en climat à fort déficit de vapeur d’eau de l’air. Pour le deuxième point, il est évident que de fortes variations temporelles du déficit de pression de vapeur d’eau (VPD) dans l’air peuvent engendrer des variations de l’efficience de transpiration et donc des incertitudes dans l’estimation de E. Il est néanmoins possible selon Abbate et al. (2004) et Kemanian et al. (2005) de corriger les valeurs journalières de ET en fonction des valeurs de VPD correspondantes afin de réduire les erreurs d’estimation.
L’intérêt primordial de l’approche illustrée à la Figure 1 est de montrer que l’efficience agronomique de l’eau consommée par une culture (EffET) dépend en premier lieu du niveau de biomasse atteint à la récolte puisque celui-ci détermine en grande partie la répartition de la consommation d’eau entre E et T. Plus la culture sera récoltée tardivement avec une biomasse élevée plus elle aura une valeur de EffET élevée. Il en résulte donc que comparer EffET entre espèces et cultivars n’a de sens qu’à équivalence de biomasse produite. Il en est de même pour les comparaisons d’efficiences entre systèmes de culture : l’irrigation ayant tendance à maintenir une surface de sol constamment humide en surface aura tendance à amplifier E par rapport à une culture non-irriguée pour laquelle l’auto-mulching du sol limitera E.
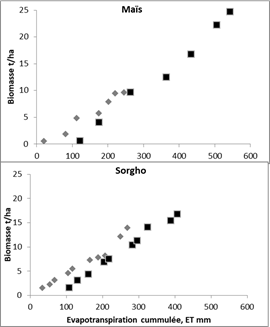
Figure 2 : Relations entre l’accumulation de Biomasse (B) et l’évapotranspiration cumulée (ET) d’une culture de maïs (a) et de sorgho (b) en sec (symboles gris) ou en irrigué (symboles noirs) :Maïs irrigué : B = 0,055 (ET-104) R2 = 0,978 ; Maïs sec : B = 0,041(ET-18) R2 = 0,973 ; Sorgho irrigué : B = 0,0504 (ET-71) R2 = 0,994 ; Sorgho sec : B = 0,046(ET-10) R2 = 0,945. (Données recalculées d’après Lemaire et al. (1999)).
La Figure 2 illustre cette approche pour le maïs et le sorgho. L’efficience agronomique de ces deux cultures (EffET) est assez similaire : 44 et 41 kg de MS/mm d’eau consommée en fin de culture respectivement en irrigué et en sec pour le maïs et 40 et 44 kg de MS/mm pour le sorgho, et elles sont voisines en sec et en irrigué malgré des différences importantes de biomasse produite et d’eau consommée. Ceci résulte en fait d’une compensation entre l’efficience de transpiration EffT et le rapport E/T. En condition irrigué le maïs a une EffT plus élevée que celle du sorgho : 55 kg de MS/mm contre 50 kg de MS/mm, mais son rapport E/T est de 0,25 contre 0,2 pour le sorgho. Pour le maïs, la sécheresse diminue très fortement EffT de 55 à 41 kg de MS/mm mais diminue aussi très fortement la part de l’évaporation du sol du fait d’une surface de sol sèche par rapport à la culture irriguée dont la surface du sol était maintenue constamment humide. Pour le sorgho, la sécheresse n’a pas affecté l’efficience de l’eau transpirée. Seule la réduction de la composante d’évaporation du sol a été réduite d’où une efficience finale supérieure à celle du maïs. Ces résultats s’accordent bien avec les observations généralement faites sur le comportement de ces deux espèces vis-à-vis de la sécheresse qui peuvent être dues à un enracinement plus dense et plus profond du sorgho par rapport au maïs (Lemaire et al. 1999) et à une sensibilité particulière du maïs au déficit hydrique au moment de la floraison. (Hall et al., 1981 ; Muchow, 1989)


