Rôles du sol
2. Rôles du sol
Dans l’alimentation hydrique des plantes, le sol va jouer deux grands rôles : d’une part, il va avoir un rôle de régulation et de stockage vis à vis des phénomènes climatiques, souvent aléatoires et irréguliers et, d’autre part, il va jouer un rôle déterminant sur la vitesse de déplacement de l'eau dans le sol et, finalement, sur la satisfaction des besoins instantanés de la plante. La grande variabilité des sols va déterminer des conditions d’alimentation hydrique très variables d’un type de sol à un autre, et même d’un point du sol à l’autre pour un même sol.
2.1 Stockage et rétention de l’eau dans le sol
Le sol est un milieu poreux, résultant d’un assemblage complexe entre particules minérales, matière organique non-vivante plus ou moins transformée et matière organique vivante. Entre ces particules minérales et organiques, ainsi qu’entre les éléments structuraux du sol, s’organise un réseau de pores connectés qui représente l’espace vide du sol qui peut se remplir d’eau et de gaz. Le volume total des vides d’un volume de sol donné est appelé porosité. Elle est de l’ordre de 50 % en moyenne, c’est-à-dire que dans 1 m3 de sol il y aura 0.5 m3 de vide, qui pourra être rempli par un mélange d’eau et d’air. Si tous ces vides étaient remplis par de l’eau (sol saturé), ce m3 de sol moyen pourrait donc contenir 500 litres d’eau[1], qui pourrait servir à l’alimentation hydrique des plantes. Cependant, un sol ne reste jamais complètement saturé en eau (heureusement, car cela créerait des problèmes d’oxygénation dans le sol et serait délétère pour les plantes). En effet, une partie de l’eau va percoler rapidement (quelques jours) sous l’effet de la gravité, dans les pores les plus gros et s’évacuer en profondeur, ou en aval d’une pente, au-delà de la zone racinaire et ne sera pas accessible aux plantes. Au bout de ce temps, l’eau ne s’évacuera plus et va être retenue dans le sol par des forces capillaires (représentant l’affinité de l’eau pour les surfaces solides et l’équilibre des forces à l’interface eau-air-solide) qui deviennent prépondérantes. La teneur en eau du sol quand ce drainage devient très faible est nommée capacité au champ (qcc) et représente la plus grande quantité d’eau disponible du sol pour la plante. A contrario, quand le sol sèche sous l’effet de la transpiration et de l’évaporation, l’eau restante dans le sol est retenue de plus en plus fortement par le solide et il arrive un moment où la plante n’est plus capable d’extraire cette eau trop liée au sol, et donc ne peut plus satisfaire la transpiration et flétrit sans possibilité de récupération. La teneur en eau correspondant à cet état est nommée point de flétrissement permanent (qfp). On considère en général que qfp correspond à un potentiel matriciel du sol de -1.5 MPa (ou -15 bars). Dans une vision classique, simplifiée, le sol pour la plante peut donc être considéré comme un réservoir d’eau avec un niveau haut, maximal, égal à qcc et un niveau minimal, où la plante ne peut plus extraire l’eau du sol, égal à qfp. On définit ainsi la réserve utile (RU) (cf. figure 1), calculée sur la profondeur maximale d’enracinement (Zr), qui correspond à la quantité d’eau disponible pour la plante par unité de surface de sol :
![]()
Exprimée en mm d’eau par mètre d’enracinement, les ordres de grandeur de la RU sont de 80-100 mm/m pour des sols sableux à 180-220 mm/m pour des sols limoneux et de 180 mm/m pour des sols argileux. L’eau n’est cependant pas disponible la même façon lorsque le stock de la RU s’épuise (l’eau étant de plus en plus retenue par le sol) et on peut définir également un point de flétrissement temporaire (qft), qui est une teneur en eau correspondant au moment où la plante ne peut plus complètement satisfaire la demande climatique par la transpiration et la régule en fermant ses stomates qui limitent les échanges gazeux au niveau des feuilles.
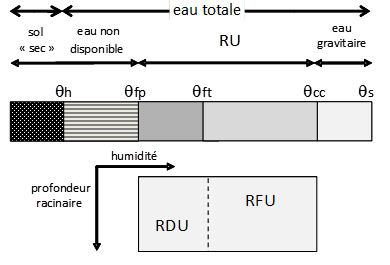
Figure 1 : Teneurs en eau caractéristiques d’un sol. qh : teneur en eau hygroscopique, sol sec en équilibre avec l’humidité de l’air ; qfp point de flétrissement permanent ; qft point de flétrissement temporaire ; qcc capacité au champ ; qs teneur en eau à saturation. Le stock d’eau entre qfp et qcc sur la profondeur racinaire définit la réserve utile (RU) séparée en une réserve facilement utilisable (RFU) et difficilement utilisable (RDU) par qft. L’eau gravitaire est l’eau pouvant circuler sous l’action de la gravité et qui est drainée en profondeur. L’eau non disponible représente la gamme de teneur en eau trop retenue par le sol pour être exploitée par la plante.
L’intérêt de cette notion de réserve utile est de pouvoir disposer rapidement d’ordres de grandeur des quantités d’eau disponibles pour l’évapotranspiration, de comparer les sols entre eux, et de pouvoir faire un diagnostic relatif à l’adéquation entre climat, culture et sol. Ce concept présente cependant des limites importantes. D’une part, la notion de capacité au champ reste imprécise (le ressuyage est un phénomène continu, dont l’arrêt est difficile à situer nettement dans le temps). Elle est parfois remplacée par des valeurs fixes précises, mais arbitraires (par exemple, la teneur en eau lorsque le potentiel matriciel du sol est égal à -0.03 MPa). D’autre part, le point de flétrissement permanent n’est pas une référence physiologique universelle (des plantes peuvent ainsi extraire de l’eau pour des potentiels inférieurs à -1.5 MPa). De plus, cette notion de réserve utile fait apparaître des notions de seuils ou des discontinuités brutales dans le domaine des mouvements de l’eau dans le sol et la plante, alors que les transferts hydriques dans le sol et l’absorption racinaire sont des phénomènes continus et progressifs. Enfin, cette approche globale en termes de quantité d’eau disponible ne fait pas apparaître le rôle central des propriétés conductrices du milieu et de l’interface sol - racines dans la satisfaction de l’alimentation hydrique des plantes. La plante y est très simplifiée (une profondeur racinaire et un point de flétrissement). Les remontées capillaires ne sont pas considérées dans cette conception du sol, alors qu’elles peuvent être non négligeables dans le cas de milieux fortement conducteurs vis à vis de l’eau et lorsqu’une nappe est située à faible profondeur. Il est alors nécessaire de dépasser l’analyse en terme de quantité globale d’eau disponible pour aborder l’analyse du transport de l’eau entre le sol et les racines en terme de flux en tenant compte des facteurs qui vont les déterminer. La dynamique de l’absorption hydrique doit être considérée ainsi que l’équilibre instantané entre l’offre du sol et la demande de la plante.
2.2 Transport de l’eau dans le sol – Notions de potentiel hydrique et de conductivité hydraulique
L’écoulement de l’eau dans le sol peut s’effectuer suivant deux grands types de mécanismes : soit de type convectif par des "chemins préférentiels" : fissures, biopores (trous de vers de terre ou de racines), espaces inter-agrégats… dans lesquels l'eau circule rapidement par gravité vers la profondeur lors d’une pluie forte (cf. ci-dessus) ; soit par le réseau de pores fins du sol, de taille micrométrique, avec des écoulements de type diffusifs, beaucoup plus lents et longs dans le temps. C’est ce second mécanisme qui est le plus fréquent et qui décrit la majorité des transferts d'eau dans le sol, notamment les transferts dans la zone d’extraction racinaire. Ce mécanisme de transport diffusif de l’eau fait intervenir l’état énergétique de l’eau dans le sol (potentiel hydrique) et les propriétés conductrices du milieu (conductivité hydraulique).
2.2.1 Notion de potentiel hydrique
De façon générale, un échange, un transport (ou une réaction) s’effectue dans le sens des énergies (représentées par un potentiel) décroissantes. L'eau du sol contient de l'énergie sous différentes formes et en quantités variées. Classiquement on distingue deux types d'énergies : cinétique et potentielle. Comme les mouvements diffusifs de l'eau dans le sol sont assez lents, l'énergie cinétique est généralement considérée comme une quantité négligeable de l'énergie totale du système (l'énergie cinétique étant proportionnelle au carré de la vitesse). C'est donc l'énergie potentielle de l'eau du sol, conditionnée par la position et l'état interne (c'est à dire en prenant en compte l'interaction avec les autres phases, les solutés ...), qui sera d'importance primordiale dans la détermination de l'état énergétique et des mouvements de la phase liquide du sol. Le potentiel hydrique (ou total) H, qui est la variable représentant l'état énergétique total de l'eau du sol, représente le travail (énergie) à fournir au sol pour en extraire l’eau. Il est la somme de la contribution des différents champs de forces (dus à la gravité, aux interactions (attraction/répulsion) avec les autres phases (le solide, l’air), à la présence de solutés ...) qui diminuent ou augmentent l'énergie potentielle de l'eau dans le sol et font que son potentiel diffère de celui de l'eau pure et libre. Par définition, les unités du potentiel sont une "énergie par unité de masse" (J.kg-1 - [L2T-2]). Pour des raisons de commodité, le potentiel hydrique peut être converti en d'autres unités. Ce peut être sous forme "d'énergie par unité de volume" en considérant que l'eau est incompressible et que sa densité ne varie pas avec le potentiel hydrique, ce qui est équivalent à une pression (Pa - [ML-1T-2]) qui est l’unité utilisée par les écophysiologistes pour le potentiel hydrique des plantes. Cette pression peut être également convertie en charge hydraulique (la hauteur d’une colonne d'eau équivalente à cette pression en m - [L]), qui est l’unité généralement employée en physique du sol et hydrologie.
Le potentiel hydrique total peut s’exprimer par la contribution des différents potentiels :
H= hm + hg + ho +…
où :
- hm est le potentiel matriciel, qui représente l’ensemble des forces liées aux interactions air - eau – solide, qui sont les forces de capillarité, d'adsorption et de tension superficielle. Dans le sol, ces forces attirent et lient l'eau sur les particules solides au point de diminuer son énergie potentielle au-dessous de celle de l'eau libre. Il faut alors fournir de l’énergie au sol pour en extraire l’eau et ce terme est négatif. Il s’annule quand le sol est saturé et devient positif quand le sol est sous une colonne d’eau (pression hydrostatique),
- hg est la résultante du travail des forces de la pesanteur,
- ho est la composante osmotique, qui représente l’effet des solutés qui abaissent l’énergie interne de l’eau. Cet effet se fait essentiellement ressentir en présence de membranes semi-perméables, c'est à dire perméables à l'eau mais pas aux solutés. C’est le cas pour des cellules végétales par exemple. Ce terme est négligé en général dans le sol.
En négligeant le potentiel osmotique, pour le sol, le potentiel hydrique total H exprimé en charge hydraulique sera simplement :
H = hm + z
Où z représente l’altitude par rapport à une référence du point considéré dans le sol (axe des z orienté vers le haut).
2.2.2 Notion de conductivité hydraulique, loi de Darcy
Le flux hydrique va donc être dirigé des zones de fort potentiel hydrique total vers les zones de plus faible potentiel hydrique. L’intensité de ce flux sera proportionnelle au gradient de potentiel total (différence du potentiel hydrique entre 2 points divisé par la distance entre ces 2 points). Cette proportionnalité entre flux et gradient de potentiel (analogue à la loi d’Ohm en électricité) est la loi de Darcy (du nom de son découvreur) en hydrologie. La constante de proportionnalité est la conductivité hydraulique (K). Cette conductivité hydraulique représente la plus ou moins grande résistance du milieu au transport de l’eau. Cette formulation de la loi de Darcy apparemment simple (proportionnalité) est en fait considérablement compliquée dans le sol par le fait que chacun des deux termes pris en compte dans cette loi (le potentiel de l'eau et la conductivité hydraulique) varie fortement, et non linéairement, en fonction des caractéristiques physiques du milieu. La conductivité hydraulique dépend ainsi des propriétés texturales du sol (sables, argiles, limons), de sa structure (continuité des pores, densité et morphologie des pores conducteurs du sol), et surtout, de sa teneur en eau ou de son potentiel matriciel. Ainsi, un sable à l'état humide est un matériau très conducteur par rapport à l'eau, tandis qu'un sol argileux est moins conducteur. Par contre, après dessèchement partiel du sol, le sable présente une très faible conductivité hydraulique généralement inférieure à celle du sol argileux. Pour un sol donné, la conductivité hydraulique diminue très rapidement lorsque le sol se dessèche : il n'est pas rare de constater des réductions d'un facteur 103 à 106 lorsqu'un sol passe de la capacité au champ au point de flétrissement permanent. Ceci montre que la possibilité de transférer l’eau du sol vers les racines peut potentiellement devenir limitante avec la chute de la teneur en eau. Pour prévoir ces phénomènes physiques non linéaires, il faut en général, avoir recours à la résolution numérique de la loi de Darcy (cf. ci-dessous).
2.3 Transfert hydrique du sol à la racine
Ainsi, si l’on considère une racine isolée, l’absorption hydrique induit le développement d’un gradient de potentiel, suivant la loi de Darcy, dans la rhizosphère sur une distance millimétrique à centimétrique. Cependant, dans un sol non saturé, lorsque la vitesse de renouvellement de l’eau à proximité de la racine est inférieure à la vitesse d’absorption de la racine, on observe une augmentation progressive de ce gradient et de la résistance aux transferts hydriques dans la rhizosphère[2]. Cette augmentation de la résistance du sol est due à plusieurs phénomènes. D’une part, les pores qui se désaturent progressivement réduisent la surface de contact racine - phase liquide. D’autre part, ce sont les pores de plus en plus petits qui deviennent progressivement les seuls fonctionnels, et ces pores sont précisément les moins conducteurs (d’après la loi de Poiseuille[3]). Ainsi, au cours de l’absorption hydrique dans un sol non saturé en voie de dessèchement, on observe une augmentation du gradient de potentiel dans la rhizosphère (ou du gradient de teneur en eau) pour vaincre la résistance croissante du sol, et assurer un flux d’absorption donné (figure 2).
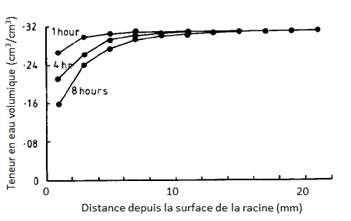
Figure 2 : Profils de teneur en eau mesuré à proximité d’une racine isolée de radis au cours du temps. (D’après Hainsworth et Aylmore, 1989)
Le transfert à l’interface sol racine va dépendre également du contact sol-racine et, concernant le sol, dépendre de la structure du sol, suivant que celui est plus moins tassé.
Dans un sol peu tassé, présentant une forte porosité et un volume de vides disponibles pour les racines important, la croissance racinaire n’est en général pas limitée par la résistance mécanique du milieu. Cependant, dans un sol peu tassé, les prélèvements d’eau (et de nutriments) par unité de longueur de racine peuvent être limités, d’une part, par le fait que si le milieu est très poreux, avec de larges pores, les zones de contact effectif entre le sol et la racine peuvent être réduites en délimitant des portions de zones où les racines sont directement en contact avec l’air, et non avec le sol ou la phase liquide. D’autre part, dans un sol peu tassé, le nombre limité de points de contacts entre les agrégats du sol va augmenter la tortuosité du milieu et réduire les surfaces d’échange vis à vis du transport de l’eau, et simultanément, diminuer la conductivité hydraulique.
Dans un sol tassé, la résistance mécanique à la pénétration opposée par le milieu au système racinaire, peut devenir un facteur limitant et ce d’autant plus que le milieu est sec, c’est à dire que sa cohésion est forte et sa plasticité faible. Cette résistance se traduit par une diminution de la croissance racinaire, qui peut s’aggraver par des problèmes d’anoxie dans ce type de milieu. Par contre, la conductivité hydraulique non saturée du sol sera augmentée par l’amélioration du contact entre les agrégats et la continuité des pores.
On peut donc concevoir qu’il existe un état structural du sol « optimal » vis à vis de l’absorption hydrominérale des systèmes racinaires, compromis entre l’intensité du contact sol - racine et le développement du système racinaire, la continuité de l’espace poral vis à vis du transport de l’eau (et des solutés). Ce type d’optimum a été mis en évidence sur des expériences contrôlées en laboratoire (Boone, 1988 ; Veen et al., 1992).
Sommaire :


