Les échanges gazeux foliaires
La feuille est à la fois un capteur de lumière et l’usine chimique où ont lieu la synthèse des sucres ainsi que d'autres molécules complexes, leur chargement dans le phloème pour redistribution vers les autres organes ou leur mise en réserve sous forme de composés à haut poids moléculaire non actifs osmotiquement comme l'amidon.
Le fonctionnement des feuilles implique des « échanges gazeux » avec l’atmosphère. En phase diurne, la photosynthèse provoque l’entrée de molécules CO2 atmosphérique et le dégagement de dioxygène O2 en volume sensiblement égal, ainsi qu’une perte en quantité beaucoup plus importante de vapeur d’eau H2O. Ces échanges gazeux diurnes sont sous la dépendance de l’état physiologique des feuilles et des conditions environnementales. La nuit, certains processus métaboliques qui restent actifs et nécessitent un apport d’énergie sont à l’origine de la respiration nocturne (absorption de dioxygène et rejet de dioxyde de carbone) et consomment une partie des assimilats synthétisés dans la journée.
Tout commence donc par l’entrée du CO2 atmosphérique gazeux dans la feuille. En présence de lumière, la machinerie photosynthétique s’active provoquant un appel de CO2 de sorte que la feuille « aspire » quasiment les molécules de CO2 présentes dans l’air.
OPTIMISATION : maximiser la photosynthèse tout en limitant les pertes d’eau
L'environnement aérien des feuilles est, du point de vue de leur fonctionnement, caractérisé par deux contraintes majeures : la très faible concentration du CO2 dans l’air (400 vpm ([1]), soit une molécule de CO2 pour 2500 molécules d’air environ) et la relative sécheresse de l'air atmosphérique par rapport à l’hygrométrie importante qui règne au sein de la feuille.
L'évolution des végétaux supérieurs a sélectionné des structures foliaires adaptées à cette double contrainte imposée par l’environnement, de façon à permettre une assimilation optimale du CO2 atmosphérique tout en limitant les pertes d'eau par transpiration. C'est ainsi que :
ñ des parenchymes lacuneux extrêmement poreux occupent une bonne partie de l'épaisseur des feuilles, facilitant la diffusion des molécules de CO2 atmosphériques et leur assimilation,
ñ l’épiderme foliaire, recouvert d'une cuticule cireuse qui limite considérablement l'évaporation, est parsemé d'une multitude de petites ouvertures microscopiques ajustables, les stomates (plusieurs centaines par mm2) à travers lesquelles s’opèrent les échanges gazeux (figure 1 et chapitre II.5).
Lorsque les stomates sont pleinement ouverts, ils n'opposent qu'une très faible résistance à la diffusion gazeuse, de sorte que la photosynthèse et la transpiration sont maximales. Par contre, lorsqu'ils sont fermés, la transpiration se limite à sa composante cuticulaire, quasiment nulle. La régulation stomatique est un phénomène très complexe où interviennent toutes sortes de facteurs : état physiologique de la plante, hormones, environnement aérien (lumière, sécheresse de l’air, teneur en CO2., etc.) ou souterrain (état hydrique du sol, etc.).
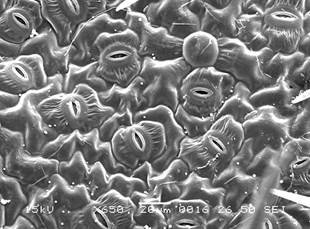
Figure 1. Microphotographie au microscope électronique à balayage d’un épiderme de Croton glanduleux. Pour montrer la disposition des stomates parmi les cellules épidermiques. Tiré de « Atlas of Croton glandulosus ».
Pour capturer au mieux les rares molécules de CO2 de l’air ambiant, les feuilles doivent avoir les stomates suffisamment ouverts, ce qui implique une perte d’eau par transpiration. L'assimilation par photosynthèse d'une seule molécule de CO2 s'accompagne de la perte de quelques centaines de molécules d'eau par transpiration, Par ailleurs, si la transpiration permet à la plante de s’alimenter en éléments minéraux provenant du sol, leur concentration dans la sève brute y est très faible, rendant nécessaire de prélever une grande quantité d’eau. Ainsi s'explique l’ampleur des besoins en eau des cultures et des arbres. Ils dépendent beaucoup du climat, de l’espèce considérée, de la structure du couvert végétal. Pour des plantes de type C3 la production d'un kilo de matière sèche nécessite plusieurs centaines de kilos d'eau. Les plantes en C4 qui présentent une adaptation anatomique particulière (figure 3) sont réputées théoriquement plus efficientes de ce point de vue, mais la question se pose de savoir comment cette adaptation se traduit in fine à l’échelle du champ et du cycle cultural complet. On a pu montrer dans certains cas que la régulation stomatique tend à optimiser l’efficience de l’eau c’est à dire à minimiser le coût hydrique de la production de matière sèche.
Mesure des échanges gazeux
Selon l’échelle considérée, la mesure des échanges gazeux (eau et CO2) fait appel à différentes techniques que nous ne ferons qu’évoquer rapidement. Les techniques de micrométéorologie s'appliquent à l’échelle du couvert végétal. A l’échelle de la plante entière ou de la feuille on utilise des chambres d’assimilation de différentes dimensions (figure 2). Il s’agit d’enceintes transparentes, contenant une ou plusieurs feuilles, un rameau, voire une plante entière, parcourues par un débit volumique d’air connu et constant Q (molair s-1) et dont on détermine la différence de concentration en CO2 et H2O entre l’entrée et la sortie. Nous avons choisi d’exprimer tous les flux en moles ce qui conduit à des notations relativement simples, où les concentrations sont exprimées en fractions molaires (voir chapitre I.2). La transpiration est ainsi donnée par :
![]() (molH2O s-1) (équat. 1)
(molH2O s-1) (équat. 1)
où ![]() et
et ![]() représentent les fractions molaires de vapeur d’eau à l’entrée et à la sortie respectivement de la chambre d’assimilation.
représentent les fractions molaires de vapeur d’eau à l’entrée et à la sortie respectivement de la chambre d’assimilation.
Avec les mêmes notations le flux d’assimilation nette de CO2 par la plante s’écrit :
![]() (molCO2 s-1) (équat. 2)
(molCO2 s-1) (équat. 2)
où ![]() et
et ![]() sont les fractions molaires de CO2 dans l’air à l’entrée et à la sortie de la chambre.
sont les fractions molaires de CO2 dans l’air à l’entrée et à la sortie de la chambre.
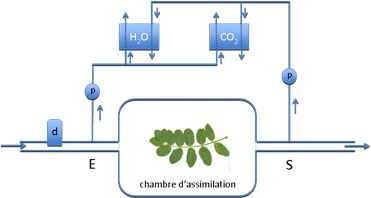
Figure 2. Schéma de principe d’une chambre d’assimilation pour la mesure des échanges gazeux des plantes. Le débit d’air qui parcourt la chambre est mesuré à l’aide du débitmètre d. Les analyseurs H20 et CO2 mesurent la différence de concentration entre l’entrée E et la sortie S de la chambre.
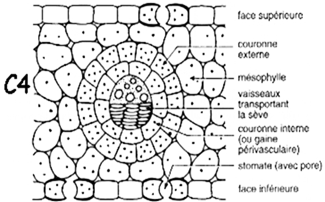
Figure 3. Coupe schématique de l’anatomie de la gaine périvasculaire chez une plante C4 (in : Photosynthesis plant C3 C4 CAM PEP carboxylase, Université d’Angers).
I.1 PHOTOSYNTHESE : les grandes lignes
La fixation du CO2 atmosphérique par les plantes est due au phénomène de photosynthèse qui d’un point de vue chimique correspond globalement à l’équation suivante :
6 CO2 + 6 H2O + énergie => C6 H12 O6 + 6 O2 (équat. 3)
Dans cette équation l’énergie est apportée par la lumière. Une partie de cette énergie est convertie sous forme chimique dans les molécules d’hydrates de carbone (= les sucres) synthétisées.
La photosynthèse est localisée dans les feuilles chlorophylliennes où l’usine photosynthétique élémentaire est le chloroplaste. On y trouve des pigments (chlorophylles, etc.) qui captent l’énergie lumineuse et la convertissent en énergie chimique sous forme de petites molécules énergétiques (ATP, NADPH) qui seront utilisées pour la réduction du CO2. Celui-ci est d’abord fixé sur le ribulose bi-phosphate (RuBP : C5H8P2O11) au site actif d’une enzyme, la ribulose bi-phosphate carboxylase-oxygénase (RuBisCO):
C5H8P2O11 + CO2 + 3 H2O => 2 C3H3PO7 + 2 H3O+ (équat. 4)
L’atome de carbone fixé par la photosynthèse est donc incorporé dans une des deux molécules C3H3PO7 (acide phosphoglycérique, APG) qui apparaissent dans l’équation 4. C’est ce qui caractérise le cycle en C3. Les molécules APG obtenues sont ensuite réduites grâce à l’ATP et au NADPH pour aboutir à la synthèse de divers composés dont notamment le glucose, sucre en C6 et le saccharose, sucre en C12 qui est le principal produit exporté par la feuille.
La RuBisCO possède par ailleurs une fonction oxygénase qui est à l’origine de la photorespiration, mécanisme qui consomme une partie des produits de la photosynthèse et conduit à une libération de CO2 en présence de lumière et d’oxygène.
Chez les plantes en C4 l’incorporation du CO2 atmosphérique conduit en premier lieu au malate ou à l’arspartate, molécules qui comportent 4 atomes de carbone. Les plantes en C4 correspondraient à une « amélioration » de la photosynthèse en C3 procurant une meilleure adaptation à la sécheresse. Elles mettent en jeu un cycle biochimique plus complexe et deux structures anatomiques spécialisées : le mésophylle et la gaine péri-vasculaire (figure 3), chacune possédant un type de chloroplastes et des enzymes particuliers. Dans le mésophylle ont lieu la photolyse de l’eau avec dégagement d’oxygène et la synthèse des composés en C4. Ceux-ci sont ensuite transportés dans la gaine péri-vasculaire où le CO2 est libéré puis incorporé dans l’acide phosphoglycérique comme chez les C3. L’avantage du cycle en C4 sur les C3, dans certaines conditions, provient de la gaine péri-vasculaire dont la structure compacte protège la RuBisCo de l’oxygène, de sorte que la photorespiration est quasiment inexistante. Ces plantes ont alors la possibilité d’effectuer la photosynthèse avec les stomates très peu ouverts, ce qui limite la transpiration.
I.2 TRANSPIRATION FOLIAIRE : origine et déterminisme
La transpiration n’est autre que le résultat de l’évaporation de l’eau des tissus foliaires et elle est soumise au même déterminisme physique que l’évaporation (cf. chapitre II.5). Elle a lieu physiquement à la surface des cellules qui tapissent l'espace poreux interne des feuilles (parenchymes lacuneux notamment). La vapeur s'échappe ensuite à travers les ouvertures stomatiques (ostioles). Rappelons que le passage de l’eau de l’état liquide à l’état vapeur nécessite un très important apport de chaleur appelé chaleur latente ou enthalpie de vaporisation de l’eau (L= 2454 kJ kg-1 à 20°C). Deux groupes de facteurs interviennent dans le déterminisme de l’évaporation d’une surface humide et peuvent dans certains cas en constituer les facteurs limitants :
1- la disponibilité de l’eau au niveau de la surface évaporante (cavitation, résistance stomatique, etc.)
2- l’apport d’énergie (chaleur) par convection, conduction et rayonnement.
L’énergie disponible au niveau des feuilles, est dissipée sous forme de chaleur sensible et de chaleur latente (évaporation) ; la part relative de l’une et de l’autre est déterminée par la résistance stomatique (théorie du bilan d’énergie, chapitre II.5).


