Modélisation des transferts couples d’eau et de carbone dans la plante
III.1 Objectifs et Hypothèses de base, principes généraux
Nous reprendrons les principes énoncés précédemment pour décrire les flux d’eau et de carbone (sucres) ainsi que leur couplage dans une plante d’architecture plus ou moins complexe. Nous avons vu l’importance de traiter séparément le xylème et le phloème mais plus généralement l’apoplasme et le symplasme des tissus. La prise en compte du carbone impose évidemment d’introduire des fonctions physiologiques dans ce modèle. On pourra trouver des exposés plus détaillés des principes du modèle utilisé ici ainsi que sur ses développements plus récents dans Daudet et al. 2002, Lacointe & Minchin, 2008.
La formulation ohmique (équat. 7, 8) a souvent été vérifiée par l’expérience, notamment sur le terrain en condition de transpiration des plantes et de potentiel hydrique négatif. Elle est d’autant mieux validée qu’elle est en accord avec la théorie (loi de Poiseuille). Nous allons largement l’utiliser dans la suite de ce chapitre pour décrire les écoulements de sève dans les vaisseaux du bois, mais également dans les tubes criblés du phloème et de façon encore plus générale pour tous les flux d’eau apoplasmique ou symplasmique qui se situent en dehors de la vascularisation sensu stricto.
Reprenons les principales hypothèses :
1. Les flux d’eau (sève brute et sève élaborée) obéissent au formalisme ohmique,
2. Ce sont les activités source et puits qui commandent la distribution des sucres dans la plante via le flux de sève élaborée.
La plante sera donc décrite comme un ensemble architecturé d’éléments hydrauliques conducteurs (tronçons élémentaires) formant un double réseau (apoplasme / symplasme) et dotés de fonctions source (S) et puits (P) de carbone (figure 8). Les tronçons élémentaires sont caractérisés par leurs résistances hydrauliques mais également par des fonctions liées aux fonctionnements hydrique (courbes pression-volume (PV), voir chapitre II.1) et carboné (fonctions source – puits), fonctions qui seront adaptées au type d’organe représenté. Le modèle devra respecter par ailleurs les bilans de carbone et d’eau. Dans l’exemple étudié plus loin l’architecture est réduite à une succession de 4 tronçons élémentaires identiques disposés en série (figure 12).
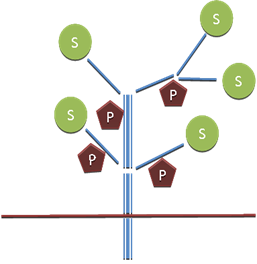
Figure 8. Structure d’un modèle architecturé. Des éléments conducteurs (tronçons élémentaires) dans lesquels circulent de l’eau (sèves) et des sucres (saccharose), sont connectés entre eux selon un schéma reproduisant la structure d’ensemble de la plante. Ces tronçons élémentaires sont dotés de caractéristiques hydrauliques (résistances, réservoirs) et de fonctions puits P et source S de carbone.
III.2 Tronçon élémentaire : contenu schématique
Nous nous appuierons sur les figures 4 et 5 pour exposer le contenu du modèle d’élément conducteur. La figure 4 montre la proximité et l’organisation spatiale des trois principaux éléments décrits dans le modèle : vaisseaux du bois, tubes criblés du phloème, parenchymes. Dans le détail toutes les plantes sont évidemment différentes mais l’organisation d’ensemble reste très comparable.
Fonctionnement hydrique : structure du sous-modèle
Du point de vue de la modélisation nous décrirons tout d’abord la structure du sous-modèle hydrique commun à tous les tronçons. La figure 9 représente schématiquement sur une coupe axiale (sagittale) schématisée, les relations hydriques existant entre les éléments anatomiques d’une structure semblable à celle de la figure 4 (coupe transversale). On a distingué l’apoplasme (partie gauche de la figure, en bleu avec notamment le xylème, parcouru par le flux de sève brute) et le symplasme (partie droite de la figure, en marron avec les tubes criblés du phloème et le flux de sève élaborée). Les parenchymes dans leur ensemble sont représentés par le gros polygone situé au centre de la figure où l’on distingue :
- l’apoplasme (ensemble des parois cellulosiques) en bleu, relié au xylème par la voie apoplasmique à travers la résistance latérale rlatapo d’une part,
- le symplasme (vacuole, cytoplasme, organites, …) en marron, relié au phloème par la voie symplasmique (plasmodesmes) à travers la résistance rlatsymp d’autre part.
La membrane plasmique figurée par un trait orange, est la frontière entre apoplasme et symplasme. C’est à son niveau que s’opère l’échange d’eau entre les deux compartiments en cas de déséquilibre.
En chaque point du réseau, les variables d’état du système hydrique sont :
Yxyl potentiel hydrique xylémien
Ypar potentiel hydrique moyen des tissus parenchymateux
Pphl pression de turgescence dans le phloème (tubes criblés)
Ppar pression de turgescence moyenne du parenchyme
P pression osmotique du parenchyme (fonction de la concentration en sucres solubles C=Csol/V, issue du sous-modèle de fonctionnement carboné)
V volume du compartiment sucres solubles ; selon les cas V sera considéré comme constant ou fonction de la pression de turgescence P.
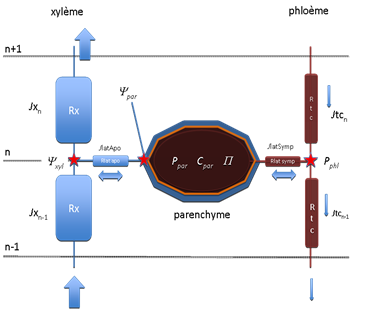
Figure 9. Structure du modèle de fonctionnement hydrique d’un tronçon conducteur élémentaire. On distingue l’apoplasme (à gauche en bleu avec le xylème, parcouru par le flux de sève brute) et le symplasme (à droite en marron avec les tubes criblés du phloème et le flux de sève élaborée). L’ensemble des parenchymes est représenté par le gros polygone central où l’on distingue l’apoplasme (en bleu) et le symplasme (en marron). La membrane plasmique figurée par un trait orange, est la frontière entre apoplasme et symplasme. On a situé les points où sont évaluées les variables d’état du système hydrique : Yxyl , Ypar , Pphl , Ppar , P (voir texte).
Le formalisme ohmique s’applique dans tous les compartiments, ce qui suppose que l’on connaisse les résistances hydrauliques axiales et transversales du symplasme et de l’apoplasme de chaque tronçon élémentaire.
Le sens de circulation des sèves s’effectue :
1. dans le sens des potentiels hydriques (Y) décroissants dans le xylème et l’apoplasme,
2. dans le sens des pressions hydrostatiques (P) décroissantes dans le phloème et le symplasme,
3. lorsque la condition d’équilibre hydrique entre symplasme et apoplasme (Y =P-P) n’est pas respectée, un échange d’eau s’établit à travers la membrane cytoplasmique. N.B. La résistance hydraulique de la membrane plasmique n’a pas été représentée sur la figure 9 mais elle est présente dans le modèle.
Outre les fonctions conductrices des tronçons élémentaires, il peut être nécessaire de considérer leur rôle de réservoir d’eau pour la plante. Ils sont alors munis de fonctions « capacitances » (analogues à des condensateurs en électricité) par l’introduction des courbes P,V (Pression, Volume) issues de données expérimentales (cf. chapitre II.1). On a représenté sur la figure 10 la relation V=f(P) utilisée dans les simulations étudiées plus loin.
Fonctionnement carboné
Le modèle de fonctionnement carboné ne concerne que le phloème et les tissus parenchymateux (figure 11). Il ne distingue pas les espèces chimiques, mais seulement trois catégories de carbone qui constituent trois variables d’état primaires :
CStruct : carbone structural inclus dans les structures stables et plus ou moins permanentes des tissus (cellulose, protéines structurales, …),
CSol : carbone soluble (glucose, saccharose, etc.). Issu de la photosynthèse ou de l’hydrolyse de Cres c’est le carbone qui circule dans le symplasme. On distingue le contenu en CSol des tubes criblés et celui du parenchyme,
CRes : carbone de réserve ou condensé, présent dans le parenchyme (sous forme d’amidon par exemple), non actif osmotiquement, mais susceptible d’être hydrolysé pour donner du C soluble.
Deux variables d’état secondaires dérivent de celles-ci connaissant le volume V des compartiments de sucres solubles :
C=CSol/V : concentration en sucres solubles,
P : pression osmotique de ce compartiment calculée par application de la loi de van ‘t Hoff détaillée au chapitre I.2 (on doit faire au préalable une hypothèse sur la nature des sucres solubles, le plus souvent le saccharose, sucre en C12).
Le bilan de carbone explicitera localement les variations de ces catégories de C, en particulier la quantité de sucres solubles CSol présents dans un tissu donné et leur concentration (C), d’où l’on déduira la pression osmotique P (cf. chapitre I.2). Les tronçons élémentaires sont dotés de fonctions source de carbone (photosynthèse, hydrolyse des réserves, etc.) ou puits de carbone décrivant leur activité physiologique (organes ou tissus en croissance par exemple). La respiration intervient dans chaque organe et l’on distinguera les respirations d’entretien RM et de croissance RG. Toutes ces fonctions sont décrites de façon plus détaillée dans la présentation d’un modèle de croissance diamétrale (§ IV.2).
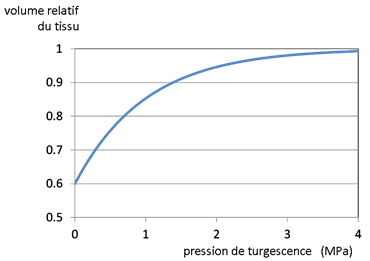
Figure 10. Relation V/Vmax=f(P) utilisée dans les simulations. Il s’agit d’un exemple non calé sur une valeur expérimentale
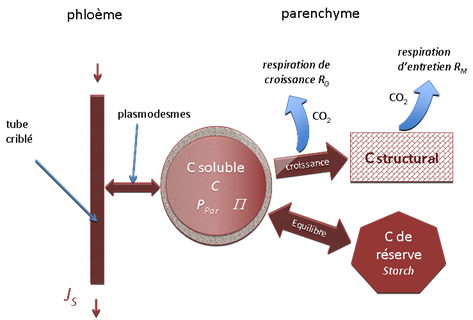
Figure 11. Schéma du sous-modèle de fonctionnement carboné d’un tronçon élémentaire. Dans les tubes criblés et les plasmodesmes, les flux d’eau symplasmique déterminés par le sous-modèle de fonctionnement hydrique (figure 9) sont associés à un flux de sucre qui fait varier le contenu et la pression osmotique P du compartiment C soluble du « parenchyme ». Ce compartiment est par ailleurs le siège d’un certain nombre de fonctions puits et sources de carbone : équilibre entre C soluble et réserves, croissance, respiration (voir texte).
Transferts couplés. Notion de couplage
Nous entendons par couplage le fait que les fonctionnements hydrique et carboné ne se conçoivent pas indépendamment l’un de l’autre. Ce couplage intervient à différents niveaux :
- l’intervention directe de la concentration en sucres solubles (CSol) issue du bilan de carbone, dans le calcul d’une composante essentielle de l’état hydrique : la pression osmotique P,
- l’entrainement des composés carbonés solubles par le flux de sève élaborée (mass flow),
- l’appartenance de plusieurs variables aux deux sous-modèles : (CSol, P),
- l’intervention simultanée de certaines variables du fonctionnement foliaire sur les flux d’eau et de carbone (ex. la résistance stomatique).
Conditions complémentaires
Conditions de conservation
Dans tous les cas il faudra respecter la condition de conservation des quantités d’eau et de carbone de façon à « boucler » correctement les bilans d’eau et de carbone. Le principe de conservation exprime que la différence entre la quantité d’un élément (eau, carbone, éléments minéraux) qui entre dans un système et celle qui en sort est égale à la variation de cet élément dans le système considéré.
Conditions initiales et aux limites
La simulation du fonctionnement de la plante placée dans des conditions particulières données suppose par ailleurs que des variables d’état aux différents nœuds (potentiel hydrique, pression de turgescence, concentration en sucre, etc.) soient initialisées ainsi que les conditions aux limites du système. La transpiration constitue une des conditions aux limites du système plante. Elle pourra soit être considérée comme une donnée, soit évaluée dans un sous-modèle de transpiration faisant intervenir l’environnement physique des feuilles et un sous-modèle de résistance stomatique. Il en va de même pour la frontière « sol » qui devra faire l’objet d’un module spécial pour déterminer l’absorption d’eau par les racines. La photosynthèse, fonction source, est une autre condition aux limites. Elle pourra être imposée arbitrairement ou faire l’objet d’un sous-modèle du même type que celui retenu pour la transpiration.


