Modèles biophysiques pour représenter le système sol-racines
5. Modèles biophysiques pour représenter le système sol-racines
5.1 Représentation des mécanismes de transport de l’eau dans le sol
Globalement, le cheminement de l’eau dans le continuum sol-plante peut être décomposé en trois étapes successives, qui vont définir les conditions de l’alimentation hydrique d’une plante :
- Au départ, la phase d'approvisionnement correspond à l’interception de l’eau du sol par le système racinaire. Cette interception se réalise par transferts d’eau dans le sol (l'eau circule dans le sol, et du sol vers les racines), mais également par la croissance du système racinaire qui permet de coloniser des zones du sol non encore exploitées ;
- L’eau passe ensuite du milieu extérieur (le sol) vers le milieu intérieur (les éléments du xylème de la racine) : c'est la phase d’absorption ;
- L’eau est alors transportée sur de longues distances dans la plante, d’abord dans les racines puis dans le système aérien jusqu’aux feuilles : c’est la phase de transport.
Ces étapes ne sont cependant pas indépendantes et une caractéristique de l’absorption de l’eau par les plantes est l’interaction constante existant entre les différents phénomènes régissant cette absorption (disposition des racines, disponibilité de l’eau, état physiologique des racines...).
Gardner (1960) a été l’un des premiers à formaliser sur des bases physiques cette vision dynamique du transport de l’eau vers la racine. Dans cette analyse (dite microscopique, i.e. à l’échelle d’un segment de racine), les racines sont considérées comme des cylindres infiniment longs verticaux (écoulement radial), prélevant de l’eau à débit constant dans un sol présentant des caractéristiques hydrodynamiques très simplifiées. Par la suite, cette approche a été étendue à divers types de conditions aux limites[4], ou à des propriétés hydrodynamiques plus réalistes ou aussi à un ensemble de racines, en supposant que celles-ci étaient identiques et réparties idéalement de façon régulière en cylindres parallèles dans le sol. Ceci revient à supposer que l’ensemble des racines extraient l’eau dans un « cylindre de sol équivalent » centré sur la racine et de rayon b égal à ![]() , Lv étant la densité volumique de racine (cm de racine / cm3 de sol). Avec ces hypothèses, une relation simplifiée (cf. Feddes et Raats, 2004) relie la chute de potentiel hydrique entre le sol et la surface de la racine et flux prélevé, en supposant que le flux d’eau est nul en b (rayon externe de sol exploité par la racine) et que le flux prélevé par la racine qr est constant (jusqu’à épuisement de l’eau du cylindre de sol) :
, Lv étant la densité volumique de racine (cm de racine / cm3 de sol). Avec ces hypothèses, une relation simplifiée (cf. Feddes et Raats, 2004) relie la chute de potentiel hydrique entre le sol et la surface de la racine et flux prélevé, en supposant que le flux d’eau est nul en b (rayon externe de sol exploité par la racine) et que le flux prélevé par la racine qr est constant (jusqu’à épuisement de l’eau du cylindre de sol) :
![]()
où ![]() est la conductivité hydraulique moyenne du sol entre les potentiels matriciels ha et hb aux rayons a (surface de la racine) et b (cylindre de sol externe prospecté).
est la conductivité hydraulique moyenne du sol entre les potentiels matriciels ha et hb aux rayons a (surface de la racine) et b (cylindre de sol externe prospecté).
Cette approche simplifiée a permis de mettre en relief un certain nombre de comportements tels que : une durée nécessaire de plusieurs jours pour extraire l’eau disponible dans un rayon de quelques centimètres autour de la racine et donc la nécessité pour la plante d’avoir un système racinaire dense pour extraire rapidement de l’eau dans un sol ; une résistance globale au transfert dans le sol liée à la conductivité hydraulique du sol et à la densité racinaire ; l’effet inverse de la conductivité hydraulique du sol et du débit sur le gradient de potentiel hydrique près des racines ; l’influence primordiale du flux qr sur la résistance au transfert dans le sol et donc la plus ou moins grande vitesse avec laquelle on atteint le point de flétrissement.
Cette approche microscopique a permis de formaliser l’effet sol sur les transferts. Cependant, la difficulté d’élargir ce formalisme analytique d’une racine à l’ensemble complexe d’un système racinaire, couplé à la sur-simplification de l’approche microscopique par rapport au système réel, occultant les problèmes d’hétérogénéité du sol et des systèmes racinaires, a nécessité une approche plus globale, débutée dans les années 80, considérant le système racinaire dans son ensemble dans le sol. Cette approche, dite macroscopique, appréhende le système racinaire comme un terme « puits » pour l’eau, distribué et diffus dans le sol, combiné à l'équation de Darcy-Richards décrivant les transferts hydriques dans le sol, pour simuler l’extraction racinaire (en une dimension, verticale ici):
![]()
Le terme S représente cette fonction « puits racinaire », exprimée en volume d'eau extrait par unité de volume de sol et par unité de temps, qui intégrée sur la profondeur d’enracinement donne l’absorption totale, c’est-à-dire la transpiration. Cette approche a été -et est toujours- très largement utilisée et une multitude de fonctions puits S ont été proposées pour décrire l’extraction de l’eau par un système racinaire. Un premier type de fonction est fondé sur des principes physiques où le flux hydrique prélevé est calculé à partir de la différence de potentiel hydrique sol - racine, pondérée par des facteurs correctifs traduisant des résistances au transfert d'eau dans le sol, vers la racine et/ou dans la plante, mais dont la formalisation et l’évaluation est difficile. Ces résistances sont largement simplifiées pour obtenir des résistances équivalentes (conductivité hydraulique du sol, résistance du transfert sol - racine obtenue en utilisant le modèle microscopique, résistance racinaire...) et/ou un terme empirique est calé, fonction de la profondeur et du temps pour rendre compte des variations de densité racinaire. Devant la nécessité de renseigner plus ou moins fortement le système sol-racine dans ce type de fonction à base physique, un deuxième type de fonction puits beaucoup plus empirique et simple utilise essentiellement des paramètres hydriques du sol, comme le potentiel matriciel. Un des formalismes les plus courants (Feddes et Raats, 2004) reprend en partie l’analogie de la réserve utile. On considère ici que l’activité des racines est maximale dans une certaine gamme de potentiel matriciel : L’extraction est considérée nulle pour des potentiels proches de zéro (anoxie racinaire) puis maximale jusqu’à un potentiel limite correspondant au point de flétrissement temporaire au-delà duquel l’extraction diminue progressivement pour être de nouveau nulle pour des potentiels du sol inférieurs au point de flétrissement permanent (cf. paragraphe 2.1). Il faut noter ici que la plante et ses mécanismes d’absorption sont représentés avec quelques paramètres « seuils » (les potentiels matriciels critiques du sol) et une densité racinaire en fonction de la profondeur.
Ces approches très globales des modèles macroscopiques, à l’échelle de l’ensemble du système racinaire, traduisent la difficulté de représenter par une boîte noire unique l’ensemble des étapes concourant à l’extraction hydrique (résistance du sol, transfert de l’eau vers la racine, absorption, transfert dans la racine) mais également le fonctionnement du système racinaire en termes de distribution dans l’espace ou d’hétérogénéité de fonctionnement. La figure 3 illustre cet effet de distribution dans l’espace des racines, où la même quantité de racines (et donc la même densité racinaire) est distribuée différemment dans un plan horizontal. La distribution régulière (Reg) est celle qui est modélisée dans les représentations microscopiques et macroscopiques des transferts, la distribution groupée (Clus) est celle qui s’approche de la réalité de la croissance racinaire d’une plante (Fig. 3A). La distribution hétérogène (Het) représente un obstacle à l’enracinement sur une partie du sol, dû par exemple à de la compaction sous des passages de roue. On constate que la distribution du potentiel matriciel du sol, au moment où la transpiration commence à chuter, est très variable suivant la distribution des racines avec de forts gradients de potentiels à proximité des racines, limitant les transferts et laissant des zones de sol peu exploitées (Fig. 3B). La conséquence est que la transpiration résultant de ces trois situations suit des évolutions temporelles très différentes (Fig. 3C), avec une durée de satisfaction de la demande climatique avant que la plante ne soit stressée qui peut être réduite de 30 à 50% par rapport à une distribution régulière de racines dans le sol. Cette différence de comportement entre les trois types de distribution des racines est due non seulement à la limitation aux transferts dans la journée, mais également à la plus faible réalimentation de la zone rhizosphérique durant la nuit avec un décrochage du potentiel matriciel à la surface des racines par rapport au potentiel moyen du sol (Fig. 3D).
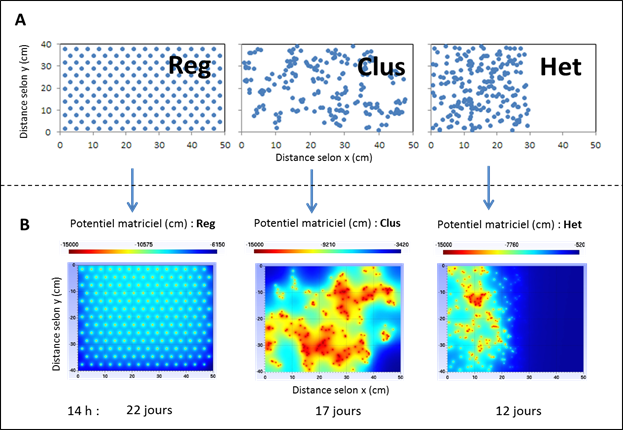
|
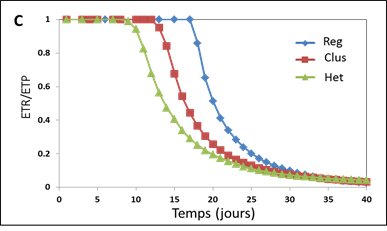
Figure 3 suite : Simulation de l’effet de la distribution des racines dans le sol sur le prélèvement hydrique.
C/ Evolution du ratio Evapotranspiration Réelle (ETR) sur Evapotranspiration Potentielle (ETP) au cours du temps pour les trois types de distribution spatiale des racines montrés en A/. On peut voir que les dispositions Clus et Het deviennent rapidement plus limitantes pour la transpiration (ETR/ETP<1) que pour Reg. D’après Beudez et al., 2013.
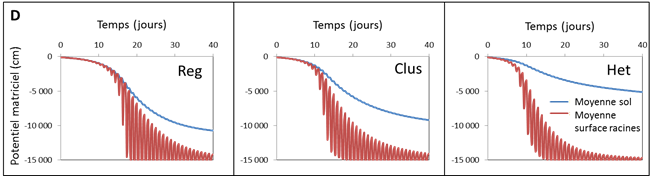
Figure 3 fin : Simulation de l’effet de la distribution des racines dans le sol sur le prélèvement hydrique.
D/ Evolution du potentiel matriciel au cours du temps, en valeur moyenne dans le sol et à la surface des racines, pour les trois types de distribution spatiale des racines montrés en A/. On peut voir des oscillations diurnes en surface des racines qui s’amplifient au cours du temps avec le dessèchement, mais aussi un décrochage du potentiel à la surface des racines par rapport à la valeur moyenne dans le sol en particulier la nuit qui signale que le sol ne parvient plus à réapprovisionner suffisamment la zone rhizosphérique la nuit pour compenser le prélèvement diurne. Cet effet est amplifié dans les cas Clus et Het. D’après Beudez et al., 2013.
5.2 Représentation de la dynamique de l’architecture racinaire
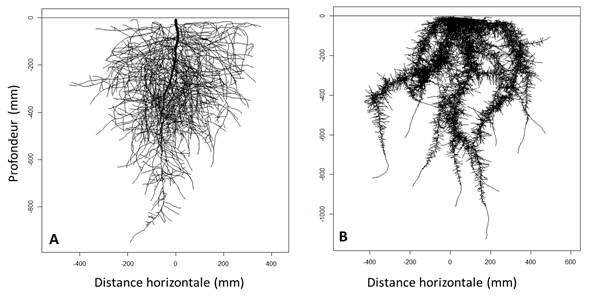
Il existe des modèles de simulation qui visent à formaliser et assembler les processus de développement racinaire pour faire des représentations dynamiques de l’architecture racinaire. Ce sont des modèles numériques dans lesquels le temps est déroulé par pas discrets (de 1 jour par exemple) et à chaque pas de temps, le système racinaire virtuel existant, représenté sous forme de petits cylindres assemblés, est modifié par l’application de processus mathématiques qui simulent les différentes formes du développement des racines que nous avons évoquées précédemment. Ainsi, certains modèles simulent conjointement l’émission de nouvelles racines d’ordre 1, l’allongement des racines existantes, leur ramification, croissance radiale, mortalité et élagage. Ces modèles de simulation peuvent également représenter des réponses du développement aux propriétés du sol, comme par exemple l’effet de couches dures qui vont limiter la croissance à une profondeur donnée, ou l’effet des températures fraiches qui vont limiter la croissance des racines de certaines espèces en profondeur. La figure 4 montre des exemples de simulations. En utilisant des jeux de paramètres différents, on arrive à représenter les différences d’architecture qui sont observées chez différentes espèces. Ces maquettes dynamiques sont utilisables pour prévoir le fonctionnement du système sol - plante, à condition d’y associer les informations qui concernent le transport de l’eau (voir ci-dessous).
|
5.3 Représentation de l’architecture hydraulique et du transfert dans le système racinaire - couplage avec les transferts dans le sol
Une des grandes limitations dans la modélisation classique (macro ou microscopique) de l’absorption est que le système racinaire est généralement peu/mal décrit, que ce soit en terme de déploiement dans l’espace et de colonisation du milieu, de croissance ou encore en terme de fonctionnement physiologique et d’interactions avec le sol et ses ressources. Une meilleure prise en compte du système racinaire est cependant nécessaire dès que les ressources du milieu commencent à devenir limitantes (plasticité, prélèvements compensatoires, régulation, interactions entre plantes) ou pour définir des idéotypes racinaires plus efficients dans l’exploitation des ressources du sol.
Le développement de modèles dynamiques d’architecture racinaire est donc une opportunité pour intégrer le déploiement et le fonctionnement racinaire d’une échelle locale (le segment de racine) à globale (l’ensemble du système racinaire) dans une représentation détaillée et interactive sol-plante de la dynamique d’absorption hydrique.
L’absorption hydrique et la transpiration sont des phénomènes passifs (sans dépense d’énergie de la part de la plante) et les flux d’eau dans ce processus vont suivre, comme dans le sol, un gradient de potentiel hydrique (du plus élevé vers le plus faible, et qui peut comporter une composante osmotique dans le cas des racines) où l’on peut distinguer deux trajets pour le transfert de l’eau au niveau des racines :
- un trajet radial de l’eau entre le milieu extérieur et les racines (le sol) jusqu’aux vaisseaux conducteurs des racines (le xylème) en traversant différents tissus cellulaires de la racine ;
- un trajet axial à l’intérieur des vaisseaux du xylème, menant l’eau des ramifications vers les racines principales et des racines principales vers les parties aériennes.
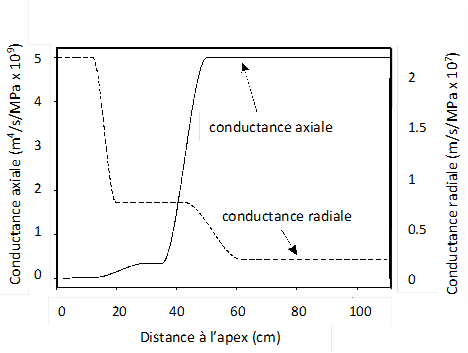
La capacité des tissus à laisser passer l’eau plus ou moins facilement selon ces trajets est caractérisée par leur conductivité hydraulique radiale (Lr , perpendiculairement à la racine) et axiale (Kh, dans les vaisseaux, dans l’axe de la racine – cf. Fig. 5). En général Lr est plus faible que Kh (on dit que la racine est anisotrope) car la résistance opposée à l’écoulement dans les parois cellulaires ou à la traversée de membranes plasmiques dans les différentes couches de tissus traversées par l’eau dans le sens radial est plus forte que dans des vaisseaux vides et fonctionnels transportant l’eau dans le sens axial. Cependant, ces conductivités hydrauliques dépendent de l’âge et de la position sur la racine. En effet, une racine poussant par sa pointe, il existe un gradient d’âge entre la pointe et la partie basale plus âgée (cf. ci-dessus) et les tissus racinaires jeunes près de l’apex vont progressivement se différencier et conférer des propriétés variables le long de la racine (cf. Fig. 5). Ainsi, le xylème est immature près de l’apex : rempli de contenu cellulaire, il ne conduit que très peu l’eau. Puis progressivement avec l’âge (ou la distance à l’apex), le contenu cellulaire meurt, les vaisseaux se vident et deviennent fonctionnels pour le transfert, en commençant par les vaisseaux les plus fins (et les moins conducteurs – protoxylème), pour finir avec les vaisseaux les plus gros (et les plus conducteurs - métaxylème). La conductance axiale va donc augmenter avec l’âge ou la distance à l’apex. Le mécanisme d’embolie, lié à la formation de bulles de gaz dans les vaisseaux de xylème, peut au contraire faire chuter drastiquement la conductivité axiale quand la plante est soumise à une sécheresse trop forte. D’autre part (voir Fig. 5), dans le sens radial, l’endoderme qui est une couche cellulaire séparant le cortex du cylindre central qui contient les vaisseaux, présente des parois cellulaires s’imprégnant de composés hydrophobes (lignine, subérine) au cours du temps. Il est une résistance majeure au transfert radial de l’eau. En effet, l’eau peut se déplacer dans le sens radial à travers les parois pectocellulosiques des cellules (voie apoplasmique) où le transfert est relativement aisé. Arrivée à l’endoderme où les parois sont plus ou moins imperméables, l’eau doit passer alors au travers des membranes plasmiques des cellules (voie symplasmique) de l’endoderme, par diffusion (osmose) ou par des canaux transmembranaires (aquaporines) et où la résistance (et le contrôle) au transfert de l’eau sont beaucoup plus forts. L’endoderme est cependant peu différencié à proximité de l’apex et cette zone présentera une conductivité radiale élevée, qui chutera avec la distance à l’apex (ou l’âge) par la subérification (terme qui désigne le dépôt de substances imperméables comme la subérine). Les aquaporines pourront moduler, dans une certaine mesure, la conductivité radiale dans le temps pour une zone donnée de la racine pour augmenter, par exemple, la capacité d’absorption du segment dans une zone plus humide du sol.
L’évolution des conductances hydrauliques racinaire variera donc fortement selon la position sur la racine et l’âge du segment avec un comportement contrasté entre conductivité radiale et axiale (figures 5 et 6). Ces variations intra-racinaires vont se dérouler également entre types de racines. Une racine secondaire ou tertiaire, de plus petit diamètre, avec également des vaisseaux de xylème plus petits et des durées de croissance courtes, se distinguera d’une racine primaire. Enfin, il y aura également des variations entre espèces, qui vont mettre en place ces variations de conductance de façon différente dans le temps. A ce titre, il faut distinguer les dicotylédones capables de croissance radiale sur les axes principaux et les monocotylédones qui ne présentent pas cette croissance. La croissance radiale, liée à une augmentation de la surface de tissus conducteurs, va permettre une très forte augmentation de la conductivité axiale, en relation avec le rôle de collecteur d’eau des racines d’ordre supérieur et de voie de transfert vers les parties aériennes.
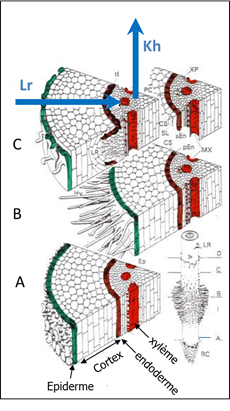
Figure 5 : Evolution des tissus le long d’une racine. Les tissus sont d’autant plus âgés et différenciés qu’ils sont localisés loin de l’apex. Les vaisseaux du xylème en A ne sont pas fonctionnels pour le transport d’eau car constitués de cellules vivantes tandis qu’en C les cellules du xylème sont mortes et les vaisseaux peuvent transporter l’eau axialement. De même l’endoderme, qui sépare le cortex du cylindre central où sont les vaisseaux, présente des parois qui s’imperméabilisent progressivement en s’éloignant de l’apex et ce faisant diminuent la conductance radiale. Lr et Kh sont les conductivités hydrauliques du trajet radial et axial de l’eau, respectivement.
|
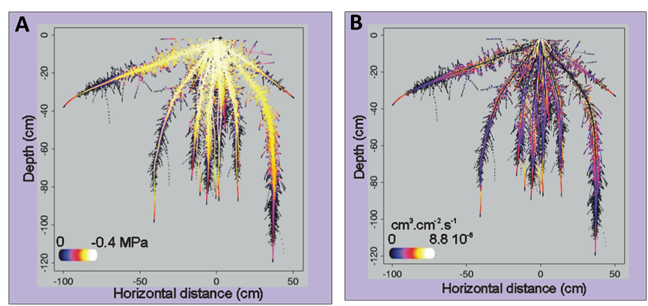
La distribution de ces conductivités hydrauliques dans le système racinaire va former l’architecture hydraulique du système racinaire. C’est cette architecture hydraulique qui va régir les interactions hydriques entre la plante et le sol : capacité d’absorption d’eau sol-plante, répartition de la demande transpiratoire sur le système racinaire, modulation du prélèvement suivant l’hétérogénéité de la disponibilité en eau du sol par interaction hydrique à travers le xylème de la plante. Ainsi, en couplant la distribution des conductivités hydrauliques des racines, fonction du temps ou de l’âge, avec un modèle d’architecture racinaire, il est possible de construire une architecture hydraulique et de simuler les flux hydriques vers et dans le système racinaire, en fonction de la transpiration et de la disponibilité de l’eau (potentiel matriciel) à l’interface sol-racine. Ce type de modèle, couplant architecture et fonctionnement est appelé modèle Structure-Fonction. La figure 7 présente un exemple de simulation d’architecture hydraulique fonctionnelle pour du maïs placé en condition de disponibilité hydrique uniforme et non limitante. On peut voir dans cet exemple que si le potentiel hydrique du xylème est relativement uniforme dans le système racinaire, sauf au niveau des parties les plus jeunes, il n’en est pas de même pour les flux d’absorption d’eau, avec des zones hétérogènes où le flux sera plus fort quand les racines latérales présentent un optimum de conductivité axiale et radiale. Ceci malgré le fait que le milieu extérieur soit homogène. L’essentiel du flux (environ 90%) est collecté par les racines latérales, tandis que les racines primaires assurent essentiellement un rôle de transport.
|
5.4 Couplage des transferts hydriques vers/dans le système racinaire avec les transferts dans le sol
Dans le sol, l’absorption hydrique et son évolution dans le temps résulteront du rapport offre (du sol) / demande (de la plante) et des interactions entre la plante (distribution de cette demande dans le système racinaire modulée par l’architecture hydraulique, réaction du système racinaire à l’hétérogénéité hydrique du sol) et le sol (transferts dans le sol proche des racines en réponse à cette demande, transferts à plus grande échelle dans le sol en réponse à l’hétérogénéité hydrique). La figure 8 illustre expérimentalement cette forte interaction système racinaire et sol pour le prélèvement hydrique. Le système racinaire a grandi dans un rhizotron, boite peu épaisse, rempli d’un sol sableux (Fig. 8A). Dans ce sol plus ou moins translucide, les variations de teneur en eau liées au prélèvement hydrique se traduisent par une plus ou moins grande transmission de lumière. Avec une caméra, on peut donc imager la distribution spatiale de l’eau dans le rhizotron au cours du temps, ainsi que la disposition des racines, (Fig. 8B et 8C). La différence des images de teneur en eau à deux dates représente l’eau extraite par le prélèvement entre ces deux instants (Fig. 8D). Sur les deux jours de transpiration de la plante, en partant d’un sol à capacité au champ, on peut voir que le prélèvement hydrique n’est pas homogène sur le système racinaire et varie au cours du temps (ce qui ne serait pas le cas en hydroponie). Ce prélèvement commence en surface, puis il se forme un front d’extraction d’eau qui se propage en profondeur et latéralement, vers les extrémités apicales du système racinaire. Ceci est le résultat de la chute de teneur en eau, due à l’absorption racinaire, qui provoque à la fois : 1) la formation de forts gradients de potentiel hydrique près des racines, 2) une chute de la conductivité hydraulique à l’interface sol-racine, qui limite fortement les transferts d’eau. En réaction à cette limitation, le système racinaire, à travers son architecture hydraulique et en particulier la distribution des conductances axiales, localise alors le prélèvement dans des zones où le sol est moins limitant. Cet exemple expérimental amplifie et accélère ce qui se passe en réalité in situ (sol peu épais, quasi 2D, sol sableux) ; les processus seront les mêmes, mais en étant plus diffus et lents dans le temps.
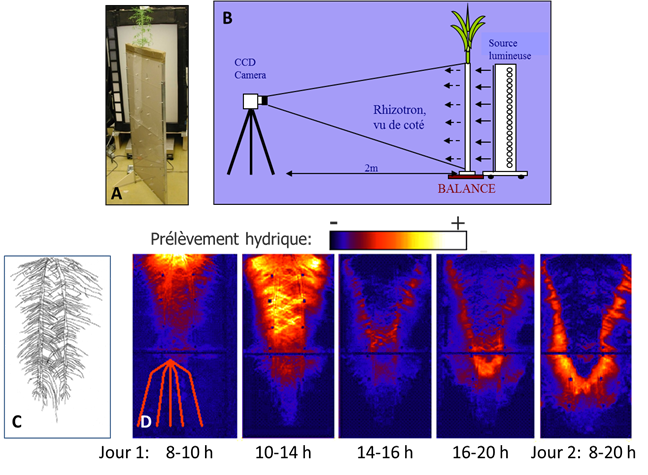
Figure 8 : Evolution au cours du temps du prélèvement hydrique dans sol sableux le long d’un système racinaire. Le système expérimental est composé d’un rhizotron (A) peu épais rempli d’un sol sableux translucide dans lequel ont poussé les racines (du lupin ici). Quand le sol sableux se dessèche suite au prélèvement par les racines, il devient plus opaque et transmet moins de lumière, ce qui donne accès à de l’imagerie de la teneur en eau dans le rhizotron au cours du temps en prenant des photos du système rétro-éclairé (B). Les différences de teneur en eau entre deux temps donnent accès au prélèvement hydrique entre ces deux moments, représentées en (D) sur 2 jours et que l’on peut rapporter au système racinaire figuré en (C) à la même échelle. Le petit schéma tracé en traits rouges indique que l’on analyse ici le fonctionnement d’un système racinaire fasciculé. La séquence diurne de transpiration se déroule de 8h à 20h et au départ le sol est à capacité au champ. On note la formation d’un front d’extraction hydrique se déplaçant au cours du temps en profondeur et latéralement sur le système racinaire, conséquence des interactions entre limitation au transfert dans le sol et réallocation du prélèvement par l’architecture hydraulique racinaire. D’après Garrigues et al., 2006.
Cette interaction non-linéaire entre le sol et le système racinaire peut être modélisée en couplant les modèles structure-fonction d’architecture hydraulique racinaire et la modélisation 3D des transferts dans le sol et à l’interface sol racine. Ceci est possible grâce à l’utilisation du potentiel hydrique, qui est la variable-clé du système sol-plante, avec les mêmes formalismes pour décrire les flux dans le sol et la plante et la continuité du processus physique dans le système sol-plante (-atmosphère). De tels modèles sont développés depuis une dizaine d’années et permettent d’étudier plus en détail les interactions entre absorption hydrique et disposition spatiale des racines, ou encore absorption et croissance, les effets de la résistance à la pénétration, les régulations biologiques (par exemples synthèse d’ABA dans les racines, modulation de la conductivité hydraulique racinaire par les aquaporines…), les effets de plasticité et de compensation du prélèvement suivant l’hétérogénéité du milieu, ou encore pour tester in silico l’efficience de prélèvement de divers génotypes d’une même espèce présentant des traits racinaires variables, dans des conditions pédoclimatiques variées et aider ainsi à la sélection de variétés (idéotypes racinaires) adaptées à diverses contraintes hydriques.
Un exemple de ce type de modélisation en relation avec l’efficacité de prélèvement est donné figure 9, pour deux génotypes de riz, l’un présentant moins de racines primaires (E-, environ 2 fois moins) émises par rapport à l’autre (E+). Les deux génotypes sont placés dans les mêmes conditions de transpiration (~ 5mm/j) et dans le même sol (limon), pour les mêmes conditions hydriques initiales (potentiel matriciel= -400 cm, soit -0.04 MPa, environ la capacité au champ). La modélisation montre qu’au bout de 7 jours de transpiration le potentiel hydrique au collet (i.e. la jonction entre racine et tige) du génotype E- (ayant moins de racines primaires) est quasiment deux fois plus négatif que le génotype E+, signalant un état de stress hydrique important. Cette différence en termes de potentiel hydrique du xylème se retrouve sur l’ensemble du système racinaire (Fig. 9A) et trouve pour partie son origine au niveau du sol et à l’interface sol-racine. On peut voir en effet (Fig. 9B) que les zones de potentiel fortement négatif dans le sol et à l’interface sol-racine sont nettement plus étendues pour le génotype E- par rapport à E+. Les flux entrant dans le système racinaire (Fig. 9C) semblent plus homogènes, à part sur la partie basale du système racinaire où ils faiblissent. Cette baisse en surface étant compensée par une augmentation en profondeur. Si le génotype E- est plus à risque vis-à-vis du déficit hydrique, le génotype E+, bien que plus fourni en racines, n’est pas réellement plus efficace dans le prélèvement de l’eau pour ces conditions car les potentiels au collet et les flux sont quasiment proportionnels à la surface racinaire des deux génotypes. Ceci est lié au fait que la conductivité radiale chez le riz semble nettement plus faible que chez d’autres céréales et apparait comme l’élément limitant des transferts dans ces conditions, par rapport au sol.
Figures 9 : Exemples de simulation par une modélisation structure-fonction du prélèvement hydrique de deux génotypes de riz où le génotype E+ est caractérisé par l’émission de deux fois plus de racines primaires que E-. Les deux génotypes sont âgés de 22 jours, dans le même sol (limon) à capacité au champ au départ et soumis à la même transpiration (5 mm/j) pendant 7 jours. (Doussan C. 2016 – non publié).
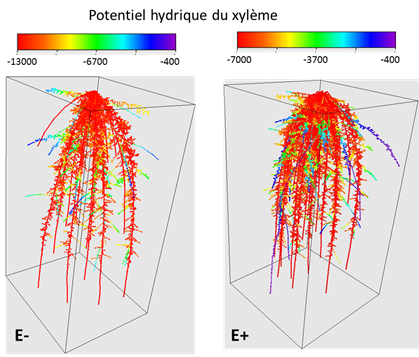
Figure 9-A : Potentiel hydrique du xylème dans le système racinaire des génotypes E- et E+. Le potentiel hydrique dans le génotype E- est deux fois plus faible que pour E+, s’approchant du seuil critique de -1.5 MPa (=-15000 cm) où la plante ne pourra plus abaisser son potentiel hydrique. (Doussan C. 2016 – non publié)

Figure 9-B : Potentiel matriciel dans le sol et à l’interface sol-racine pour les génotypes E- et E+. Les zones de potentiel matriciel faible (dans sol et à l’interface sol-racine) sont plus étendues pour le génotype E-. En moyenne, le potentiel à l’interface sol racine du génotype E- est plus négatif de -1500 cm par rapport au génotype E+. (Doussan C. 2016 – non publié)

Figure 9-C : Distribution des (densités) de flux d’eau entrant dans les racines pour les génotypes E- et E+. E- extrait à peu près deux fois plus d’eau par unité de surface de racine que E+, avec un patron spatial presque similaire. La diminution en surface de prélèvement, plus prononcée pour E+, est compensée par une augmentation des flux en profondeur. (Doussan C. 2016 – non publié)
Si ce type de modèle structure – fonction est à même d’intégrer de façon détaillée différents processus sol et plante, d’être couplé avec d’autres processus comme l’absorption/transfert de nutriments et d’intégrer des processus spatiaux de l’échelle du segment de racine (le millimètre) à l’échelle d’un système racinaire (le mètre), il n’en reste pas moins que ce type de modélisation est très exigeant en paramètres sol et plante, pas toujours facilement accessibles, assez difficile numériquement et pouvant être long en temps de calculs. Tous les processus sol-plante et leurs interactions ne peuvent être représentés dans ce cadre avec le même degré de détails et, comme dans les autres types de modélisation, il est nécessaire alors de faire des compromis entre détail, efficacité et réalisme.
Sommaire :


