Exemples de simulation
Transpiration des arbres
Nous prendrons l’exemple d’un jeune pin maritime poussant en plantation dans le sud-ouest de la France. Le sol est colonisé par les racines des arbres jusqu’à une profondeur de 1 m environ, lui offrant 120 mm de réserve utile. Cette valeur est assez faible, mais courante sur les sols sableux des Landes de Gascogne. L’ensemble des arbres de la plantation (1500/ha) a un indice foliaire (LAI) égal à 3,2, ce qui est faible, mais qui correspond à la réalité. Chaque arbre a en moyenne une surface d’aiguilles développée d’environ 20 m². Concernant les conditions climatiques, nous considèrerons ici deux successions de 33 journées sans pluie : d’abord une série théorique de belles journées estivales identiques puis une seconde série de mesures réelles. La figure 8 représente l’évolution de l’ETP et de la transpiration au cours de ces deux périodes.

Figure 8. A : simulation de la transpiration pour une succession de 33 journées à ETP identiques (journées ensoleillées en été), valeurs semi- horaires. B : idem avec des données météorologiques observées sur le site.
On constate sur la figure 8A une diminution progressive de la transpiration des arbres, en réponse au dessèchement du sol, celle-ci s’annulant presque au bout des 33 journées. Ce phénomène est moins marqué sur la figure 8B, en raison de la plus faible demande transpiratoire (ETP) en particulier lors des journées couvertes (voir les journées 12 à 18).
Potentiels hydriques
Connaissant la transpiration et les paramètres hydrauliques des arbres, nous pouvons maintenant calculer :
- la diminution de la réserve en eau disponible dans le sol, donc le potentiel hydrique de celui-ci ;
- le potentiel hydrique dans les feuilles (les aiguilles) de l’arbre ainsi que dans différents points du trajet de la sève brute. Nous représentons sur la figure 9 les potentiels hydriques dans le sol, le tronc et les feuilles.
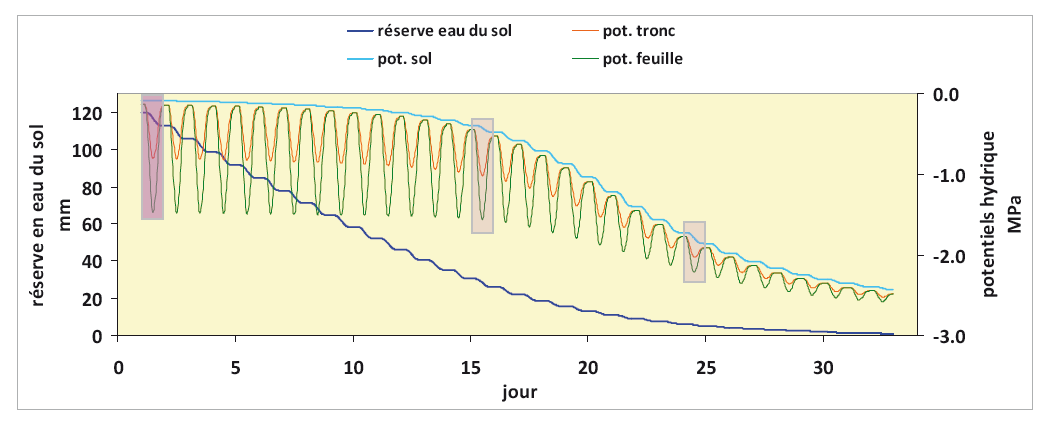
Figure 9. Simulation de l’évolution, au cours de la période d’étude, de la réserve en eau du sol (échelle de gauche) et des potentiels hydriques à différents niveaux du système sol-plante-atmosphère (échelle de droite) : le sol, le xylème du tronc et les feuilles. Les 3 journées représentées sur la figure MOD10 ci-dessous sont entourées avec des rectangles roses.
Pendant les 10 premiers jours, les potentiels hydriques simulés de l’arbre sont comparables d’un jour à l’autre, bien que la réserve en eau du sol diminue. Ceci est du au fait que le potentiel hydrique du sol ne diminue que faiblement au début d’un dessèchement (voir Fig. 5). Ensuite, lorsque le dessèchement du sol se poursuit, les potentiels hydriques, diurnes et nocturnes, diminuent à tous les niveaux du SPAC. La figure 10 ci-dessous présente trois des journées de cette cinétique : au début de la période, lorsque l’eau du sol n’est pas limitante, une journée où la sécheresse est moyenne et enfin une journée au plus fort de la sécheresse.
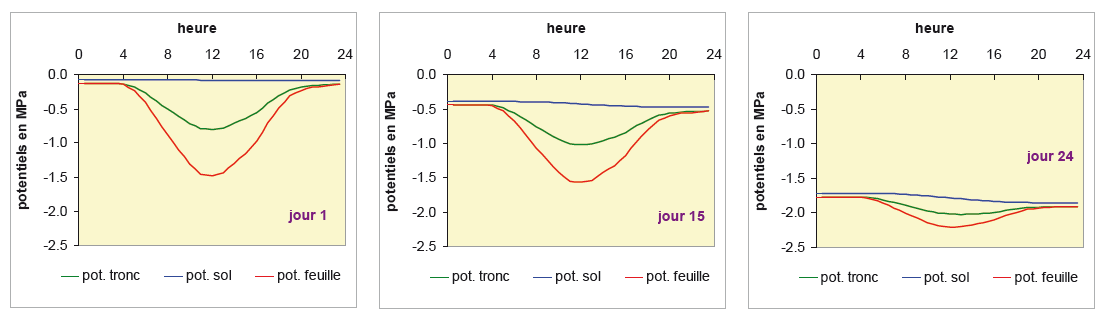
On voit clairement entre ces 3 journées la diminution du potentiel hydrique du sol. Pour chaque journée, la valeur de ce potentiel est légèrement plus faible le soir que le matin, en raison de la consommation en eau par l’arbre au cours de la journée. Les potentiels hydriques dans le tronc et dans les feuilles suivent une course sensiblement parallèle, mais on note au fur et à mesure du dessèchement un aplatissement et un rapprochement des deux courbes, en raison de la régulation stomatique qui réduit la transpiration. Au jour 24, on constate que le potentiel foliaire minimum (environ -2.2 MPa) devient proche du potentiel de base (la valeur atteinte en fin de nuit, ici d’environ -1.8 MPa), conséquence d’une fermeture stomatique presque totale.
Des comportements hydriques différents selon les espèces lors d’une sécheresse : les facteurs de variation
Dans une situation de stress hydrique, la cinétique de diminution du potentiel hydrique d’une plante est particulièrement importante à connaître pour être capable de prédire sa réponse, car le potentiel hydrique de ses organes impacte des fonctions essentielles, notamment sa photosynthèse et sa croissance en longueur et en diamètre, mais aussi sa résistance aux attaques biotiques (insectes et champignons).
Deux paramètres modulent principalement cette cinétique du potentiel hydrique : 1) la réponse des stomates, variable selon les espèces, 2) la réponse de la conductance hydraulique des tissus du xylème avec, comme composante déterminante, la vulnérabilité à la cavitation, qui diffère selon les espèces. Nous donnons ci-dessous quelques exemples illustrant les conséquences des variations de chacune de ces deux composantes au cours de cinétiques de dessèchement.
La régulation stomatique
De façon assez générale, les stomates se ferment lorsque la contrainte hydrique dans le sol augmente (voir chapitre II.5), mais la précocité de cette fermeture est variable selon les espèces.
Certaines espèces sont caractérisées par une fermeture stomatique précoce et efficace. La figure 11A montre les variations du potentiel hydrique foliaire d’une telle espèce au cours d’un dessèchement, en partant d’un sol bien hydraté. Cette espèce, par exemple un pin maritime ou un hêtre, se caractérise par une forte transpiration au début de la période, provocant une chute rapide de la réserve en eau du sol (ici à partir du jour 10) et par conséquent de son potentiel hydrique. A un certain niveau de dessèchement, la fermeture stomatique devient presque totale, ce qui conduit à une forte réduction de l’amplitude des oscillations jour-nuit (au-delà du jour 20). Le potentiel hydrique du sol et des feuilles se stabilise alors. Ce comportement, qui limite la diminution du potentiel hydrique dans les organes de la plante est qualifié d’isohydrique.
La réponse d’une espèce à fermeture stomatique tardive est très différente (Fig. 11B). On constate que la diminution du potentiel hydrique des feuilles est plus progressive. On voit aussi que l’amplitude des variations jour-nuit se maintient plus longtemps que dans l’exemple précédent. Ces plantes sont appelées anisohydriques, ce sont des plantes qui n’assurent pas le maintien de leur potentiel hydrique au-dessus d’un certain seuil. C’est par exemple le cas de la betterave dont les feuilles flétrissent en période de sécheresse, signe de perte de turgescence (voir chapitre I.2) des cellules foliaires.
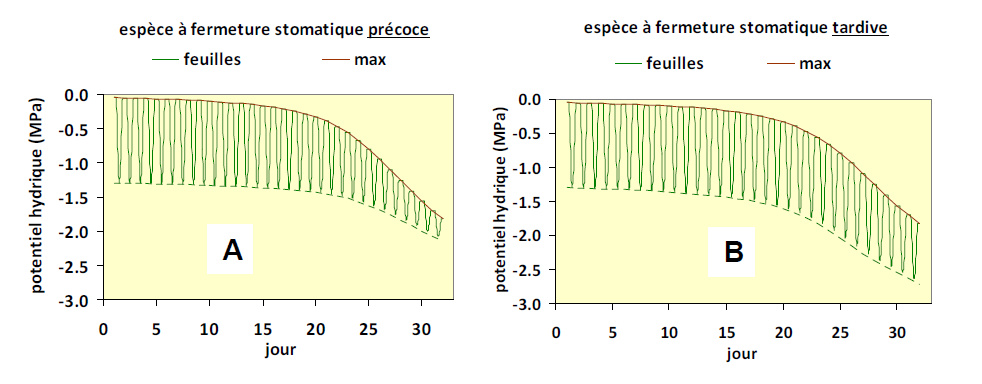
Figure 11. Evolution du potentiel hydrique foliaire au cours d’un dessèchement du sol, en partant de sa capacité au champ au premier jour. On a représenté les enveloppes supérieure (= la courbe de couleur brune suivie par les potentiels de base, qui sont très proches du potentiel hydrique du sol) et inférieure (= les potentiels hydriques minimums, atteints en milieu de journée). Deux types contrastés sont représentés. Figure A : cas d’une espèce à fermeture stomatique précoce ; figure B : cas d’une espèce à fermeture stomatique tardive.
On parle de stratégies développées par les plantes face à la sécheresse, et les deux comportements contrastés décrits ci-dessus présentent chacun leurs avantages et leurs inconvénients.
Les espèces du premier type, appelées espèces évitantes, présentent un bon contrôle du dessèchement et se protègent rapidement lors de longues périodes sans pluies ; elles évitent ainsi de dépasser des seuils irréversibles portant atteinte à leur tissus. Toutefois, elles ne fonctionnent (transpiration, photosynthèse, croissance…) que sur des durées assez courtes. Les espèces du second type, appelées espèces tolérantes, ont la possibilité de fonctionner plus longtemps lors d’une séquence sans pluies. Mais il y a pour ces espèces le risque de dégradation de l’intégrité de leurs fonctions et de leurs tissus. Pour y faire face, dans des conditions de dessèchement prolongé et intense, ces plantes doivent être capables :
· soit de survivre et de protéger leur intégrité grâce à des adaptations physiologiques, comme chez beaucoup d’espèces méditerranéennes ou subtropicales,
· ou bien d’adopter une stratégie tout à fait différente : la fanaison suivie de la mort de la partie aérienne, seule la partie racinaire survivant alors dans le sol, ou bien il se produit une stimulation de sa fructification, généralement suivie de la mort de la plante.
La conductance hydraulique
D’une manière générale, la conductance hydraulique de la plante diminue avec l’augmentation de la contrainte hydrique, souvent pour plusieurs raisons, principalement la dégradation de son continuum sol-racines et celle de sa capacité de conduction de l’eau dans son xylème consécutive à la succession cavitation-embolie (voir chapitre II.7).
Tout comme pour la régulation stomatique, la diminution de conductance hydraulique diffère selon les espèces et affecte l’évolution des potentiels hydriques en conditions de dessèchement. Nous en donnons ici un exemple pour deux plantes, dont les conductances hydrauliques totales présentent des réponses différentes lors de l’abaissement du potentiel hydrique du sol (Fig. 12A). Ces deux espèces montrent au début du dessèchement du sol une diminution rapide de leur conductance hydraulique, puis au-dessous de -0.5 MPa environ, cette diminution est plus importante chez l’espèce 2, dont le système conducteur se dégrade par les mécanismes de cavitation et d’embolie. Les conséquences sur les potentiels hydriques sont présentées sur les figures 12B1 et 12B2.
On constate sur cette figure qu’à partir du jour 20, la différence la plus visible entre les deux espèces est le niveau atteint par les potentiels hydriques minimums, qui devient nettement plus faible pour la plante de type 2 (-2.6 MPa) que pour celle de type 1 (-2.0 MPa).
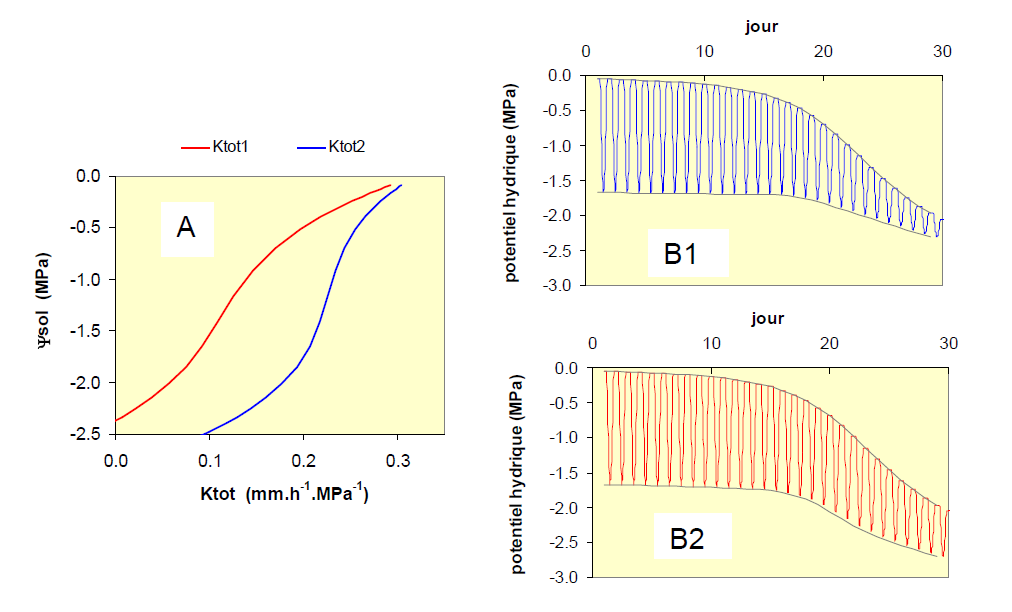
Figure 12. A : cas de deux plantes présentant des réponses contrastées de leur conductance hydraulique totale Ktot (entre le sol et les feuilles) à la contrainte hydrique du sol, exprimée ici au moyen du potentiel hydrique du sol. La plante de type 1 (courbe bleue) montre une bonne stabilité de Ktot, même lorsque la contrainte hydrique est forte, alors que la plante de type 2 (en rouge) voit sa conductance hydraulique chuter régulièrement avec l’augmentation de la contrainte, c'est-à-dire lorsque le potentiel du sol diminue. Figures B : Conséquences sur l’évolution des potentiels hydriques de ces deux propriétés, avec le tracé des limites supérieures et inférieures du potentiel hydrique foliaire, B1 correspondant au type 1 et B2 au type 2.
Le volume des réservoirs hydriques internes
Il est intuitif de penser que le volume des réservoirs pouvant échanger de l’eau avec le flux de transpiration – se dessécher et se réhydrater (voir chapitre II.3) – a des répercussions sur la dynamique des flux d’eau dans la plante. Le modèle présenté ici permet d’en voir quelques conséquences sur les cinétiques diurnes de transpiration et d’absorption. Nous prendrons dans la figure 13 l’exemple de deux plantes présentant une différence marquée du volume d’eau échangeable (leur capacitance hydraulique).
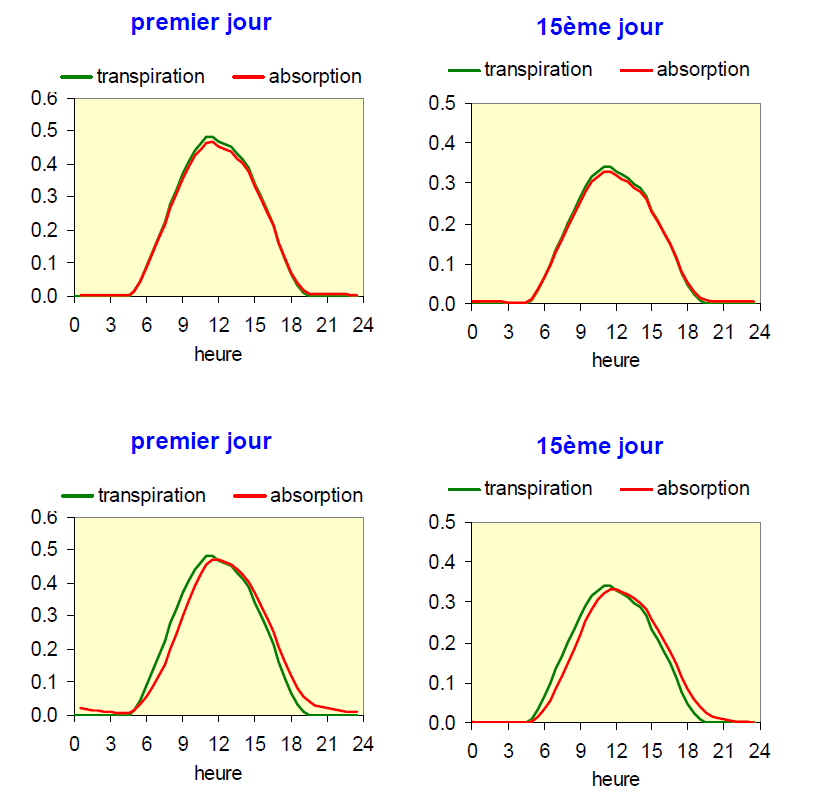
Figure 13. Simulation de la transpiration et de l’absorption d’eau du sol pour deux plantes caractérisées par des réservoirs d’eau échangeables de dimensions différentes. En haut : cas d’un réservoir dont le volume est limité ; en bas : avec un réservoir de volume important. On a représenté la première journée, où le sol est humide et la quinzième journée de dessèchement.
Un sous-modèle décrivant les variations de diamètre du tronc d’un arbre
Nous décrivons ici un petit modèle, qui est connecté au modèle plus général décrit ci-dessus ; pour cette raison, il est appelé sous-modèle .
Les variations diurnes de dimensions d’un organe végétal non totalement rigide (voir des exemples donnés au Chapitre II.3 pour des fruits, des feuilles ou des tiges) reflètent celles de sa teneur en eau, directement dépendante des variations du potentiel hydrique dans le xylème connecté à cet organe. Néanmoins, au cours d’une journée, on observe souvent un décalage, parfois important, entre les variations de la demande climatique (ETP) celles du diamètre de l’organe. Ce phénomène est illustré sur la figure 14 : alors que le maximum d’ETP intervient vers 12 h (heure solaire), le rayon du tronc du pin maritime étudié continue de se rétracter et n’atteint sa valeur minimale qu’à 15 heures, soit 3 heures plus tard. Enfin, lorsque l’arbre est en croissance, le rayon de son tronc à l’issue de ce cycle de 24 heures a une valeur supérieure à celui de la veille.
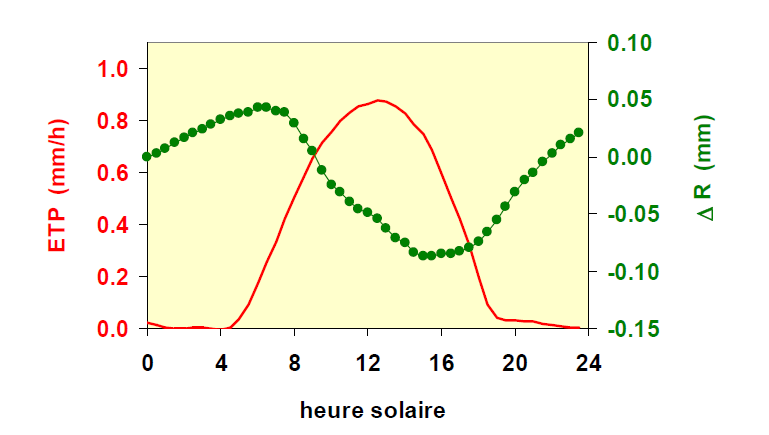
Figure 14. Variation, au cours d’une belle journée estivale, de l’ETP (en rouge, échelle de gauche) et du rayon d’un jeune pin maritime (en vert, échelle de droite). On a décalé arbitrairement la mesure du rayon de l’arbre pour qu’elle soit à zéro au début de la journée. Le rayon augmente un peu en fin de journée et la nuit puis diminue dans la journée pour atteindre son minimum vers 15 heures.
Nous décrivons ci-dessous la chronologie des mécanismes mis en jeu :
· Nous supposons d’abord qu’en fin de nuit, tous les tissus de l’arbre sont en équilibre hydrique entre eux ; il n’y a pas de mouvements d’eau, hormis éventuellement ceux liés à la croissance de certains organes. Le potentiel hydrique de l’ensemble des tissus du tronc est stabilisé à une valeur Yr ;
· Le matin, à partir du lever du soleil, l’ETP augmente, induisant la mise en route de la transpiration, provoquant la diminution des potentiels hydriques dans toute la plante. La valeur des potentiels hydriques est en relation directe avec le niveau de transpiration et les conductances hydrauliques des organes conducteurs ;
· Intéressons-nous maintenant aux tissus corticaux du tronc (le phloème et la zone génératrice du phelloderme, qui gainent le xylème ; voir Chapitres II.2 et II.3) : l’écart de potentiel hydrique entre le xylème Yx et celui des tissus considérés (Yr) augmente. Cet écart de potentiel provoque un flux d’eau allant des tissus corticaux vers le xylème, ce flux étant d’autant plus grand que l’écart entre Yr est Yx l’est. Ainsi, ces tissus se vident et viennent supplémenter la transpiration. Leur volume diminue et, comme ils sont partiellement élastiques, leurs dimensions, notamment leur épaisseur, diminue elle aussi. Le tissu ayant perdu de l’eau se trouve alors porté à un niveau Yr plus faible que précédemment. Ce mécanisme se poursuit tant que Yx reste plus faible que Yr. Lorsque l’ETP et en conséquence la transpiration décroissent en fin de journée, ce mécanisme s’inverse : le potentiel Yr est plus faible que Yx, ce qui provoque un flux d’eau allant du xylème au tissu concerné. Celui-ci se remplit progressivement, parfois jusque tard dans la nuit, jusqu’à que l’équilibre entre les potentiels hydrique soit rétabli. Les variations du niveau de remplissage des tissus corticaux sont ainsi en retard sur la contrainte imposée par le climat (ETP), comme on peut le voir sur la figure 14.
Sommaire :


