Le continuum sol-plante-atmosphère
3. Le continuum sol-plante-atmosphère
Du point de vue des transferts hydriques, le sol la plante et l'atmosphère forment un continuum c’est-à-dire un ensemble de compartiments qui communiquent entre eux : l’eau du sol transite par les racines à travers la plante jusqu'aux feuilles et les autres organes transpirants (bourgeons, fleurs, fruits...) avant de rejoindre l'atmosphère. Cet ensemble de trois éléments forme ce qui est appelé le continuum sol-plante-atmosphère, ou CSPA en abrégé. La plante est ainsi un élément biologique reliant deux compartiments physiques, le sol et l’atmosphère.
3.1. Lois circulation dans le CSPA et analogie électrique
Contrairement aux organismes qui vivent dans une eau liquide, très facilement accessible, la plante terrestre se heurte à une difficulté majeure, celle d’extraire l’eau du sol. Cette eau adhère aux particules de sol, matière organique, sables, argiles et limons, parfois très fortement lorsqu’il y a une sécheresse. Pour fonctionner et croître, la plante doit être capable d’aspirer cette eau avec d’autant plus de force (ou d’énergie) que le sol est desséché ; ceci sera développé ci-dessous.
Le mécanisme qui permet à la plante d’aspirer l’eau du sol est relativement simple et connu depuis plus d’un siècle (Dixon, 1894, 1914). Pour décrire correctement le déterminisme de ce phénomène, il faut partir des feuilles et non pas des racines : en effet, l’eau est aspirée par les feuilles et non pas poussée par les racines[3]. Le mouvement de l’eau se transmet en cascade dans les pétioles, les rameaux, branches, tige jusqu’aux racines, par lesquelles pénètre l’eau du sol.
Le concept de CSPA a été proposé par Huber (1924), puis développé par Gradmann (1928) et Van den Honert (1948). Le flux d’eau qui traverse les différents compartiments du CSPA existe du fait de l’existence de différences de potentiel hydrique entre eux. Le potentiel hydrique est une grandeur physique, qui exprime l’état de liaison de l’eau, aussi bien dans un organe végétal, que dans le sol ou l’atmosphère. Le potentiel hydrique de l’eau libre, comme celle d’un lac, est égal à zéro. Par convention des physiciens, lorsque l’eau se trouve sous pression, son potentiel hydrique est positif ; c’est le cas de l’eau de nos canalisations domestiques. Mais, dans la plante, le potentiel hydrique est presque toujours négatif. Cela signifie que l’eau est retenue dans la plante, en état de dépression. On peut faire la comparaison avec une éponge (pas trop humide) : son potentiel hydrique est négatif et il la faut presser, donc exercer une force, pour en extraire l’eau. L’unité du potentiel hydrique est le Pascal (Pa)[4]. Dans une plante qui transpire, le potentiel hydrique est de plus en plus négatif sur le trajet allant du sol aux racines, puis aux tiges et aux feuilles. Ce potentiel est ainsi très négatif dans les feuilles et il l’est encore plus dans l’atmosphère où il peut être inférieur à – 100 MPa. Lors de belles journées estivales, le potentiel hydrique des feuilles descend à des valeurs d’environ – 1 MPa chez les plantes herbacées ou les cultures, et jusqu’à – 2 MPa voire au-delà chez les arbres.
Les chercheurs mesurent le potentiel hydrique des feuilles ou des rameaux feuillés grâce à un dispositif appelé chambre à pression (photo 2; voir aussi Chap. II.3). Il s’agit d’un appareil simple : la feuille est enfermée dans une petite chambre en métal épais, son pétiole dépassant au-travers d’un joint élastique. Un gaz sous pression est injecté lentement pendant que l’opérateur regarde l’extrémité de ce pétiole avec une loupe. Lorsque la sève brute commence à sortir, il note cette pression, par exemple 1.2 MPa : c’est celle qui contrebalance le potentiel hydrique de la feuille, donc ici -1.2 MPa.
En conditions de bonne alimentation en eau, c’est-à-dire lorsque le sol est humide et que les stomates foliaires sont ouverts, l’eau transite du sol, où elle est à un potentiel « haut » c’est-à-dire, faiblement négatif (de l’ordre de quelques fractions de MPa), vers l’atmosphère, où le potentiel hydrique de la vapeur d’eau de l’air est bien plus négatif (plusieurs dizaines de MPa). Nous approfondirons plus loin dans ce chapitre les liens entre potentiel hydrique et flux d’eau dans la plante.

Photo 2. La chambre à pression, appareil de mesure du potentiel hydrique des feuilles. A gauche, la feuille est enfermée dans un sachet en plastique afin de bloquer la transpiration et son pétiole est inséré dans le couvercle de la chambre, l’étanchéité étant assurée par un joint en caoutchouc. A droite, la pression à laquelle la sève émerge du pétiole de la feuille est équivalente à l’opposé du potentiel hydrique de l’eau dans la feuille
Depuis longtemps les chercheurs ont établi le parallèle entre la liaison fonctionnelle reliant flux d’eau et potentiels hydriques de la plante et la circulation d’un courant dans un circuit électrique. Ce parallèle, l’analogie électrique, a été proposé dès 1948 par des précurseurs des recherches sur le fonctionnement hydraulique des plantes comme Van den Honert. Au centre de cette analogie, la loi d’Ohm, bien connue, relie de façon proportionnelle le courant dans un circuit électrique à une différence de potentiel électrique entre ses bornes. Le facteur de proportionnalité est la résistance électrique. Dans le cas d’une plante, on écrit que le flux (ou débit) d’eau est proportionnel à la différence de potentiel hydrique entre ses deux extrémités, constituées par les feuilles et les racines. Cette loi s’applique de la même façon à un organe, une racine ou la tige par exemple, voire à un segment de cet organe. La résistance à la circulation de l’eau dans la plante est appelée résistance hydraulique (son inverse s’appelle conductance hydraulique).
3.2. Les différents régimes de transfert d’eau dans la plante
On distingue différents régimes de transferts hydriques dans la plante selon que les flux sont constants ou variables sur un intervalle de temps et lorsque l’absorption de l’eau par les racines et la transpiration foliaire sont ou non égales. Des flux d’eau qui ne varient pas dans le temps sont qualifiés de régimes permanents. Dans le cas où il y existe des variations dans le temps, ces flux sont en régime variables. Lorsque transpiration et absorption sont égales, le flux est qualifié de conservatif ; dans le cas contraire, le régime de flux est non conservatif. Ces différents types de transfert sont schématisés sur la figure 1.
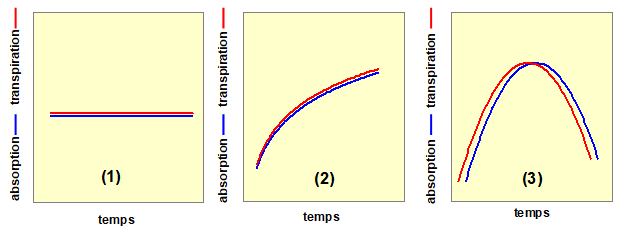
Figure 1. Les différents types de transfert hydrique dans une plante, définis en comparant la transpiration foliaire et l’absorption de l’eau par les racines. En (1), la transpiration et l’absorption ne varient pas dans le temps, le régime des flux est permanent. De plus, comme à tout moment absorption et transpirations sont égales, le flux est conservatif. En (2), le flux est conservatif mais le régime est qualifié de variable. En (3), le flux est variable et non conservatif, car absorption et transpiration ne sont pas égales (sauf juste au moment où les deux courbes se croisent).
Nous verrons, dans les exemples présentés plus loin que la plante, en conditions naturelles, fonctionne rarement en régime permanent, car la demande transpiratoire, d’origine climatique, varie au cours de la journée (course du soleil, nuages, pluie, etc.).
3.3 . Les modèles de circulation de l’eau et quelques exemples
Pour une plante entière, le modèle de fonctionnement hydrique le plus simple est représenté sur la figure 2. Sur ce schéma, la différence de potentiel s’applique entre le sol, plus précisément au niveau de l’interface entre les particules de sol et les racines fines, et les feuilles.
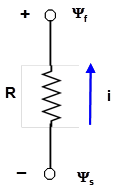
Figure 2. Le modèle le plus simple pour décrire le fonctionnement hydrique d’une plante est basé sur l’analogie électrique : R représente la résistance hydraulique au transfert de l’eau, i le flux d’eau, Yf et Ys les potentiels hydriques, respectivement au niveau des feuilles et du sol.
Des mesures simultanées de flux et de potentiel hydrique dans une plante montrent la pertinence d’un tel modèle. La figure 3 représente, pour un arbre (ici une espèce tropicale en Guyane dans une plantation), les variations au cours d’une journée du flux de sève brute mesuré dans le tronc et celles du potentiel hydrique foliaire, mesuré au moyen d’une chambre à pression. Le potentiel hydrique des feuilles diminue dès le lever du soleil pour attendre une valeur minimale en milieu de journée pour remonter ensuite. On peut observer l’étroit parallélisme entre la transpiration et le potentiel hydrique des feuilles : la relation entre le flux de sève et le potentiel hydrique est linéaire. Le potentiel hydrique stable atteint en fin de nuit (en effet, lorsque le sol est moyennement à très desséché, la plante met souvent toute la nuit pour atteindre une valeur stable) est appelée potentiel de base, qui est souvent légèrement inférieur au potentiel hydrique du sol. C’est un indicateur précieux et fiable des conditions d’alimentation en eau du sol, qui fait couramment l’objet de mesure par les chercheurs. On note aussi sur cette figure que le potentiel hydrique de base ne remonte pas à zéro pendant la nuit. Il est en effet rare que la plante soit totalement hydratée, même lorsque le sol est humide.
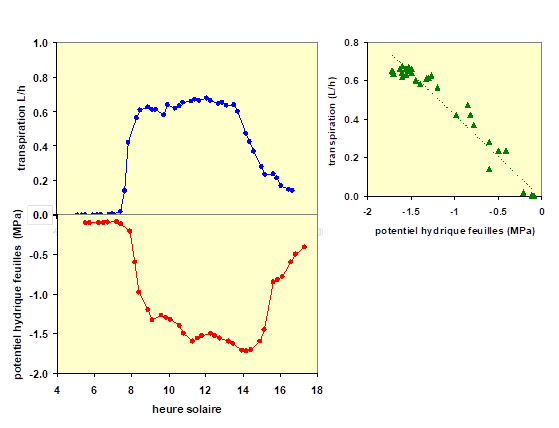
Figure 3. Transpiration et potentiel hydrique d’une plante sont étroitement liés : le matin, l'augmentation de la transpiration (courbe en bleu en haut à gauche) provoque la diminution du potentiel hydrique (courbe en rouge, en bas à gauche) dans l'arbre. La relation entre transpiration et potentiel hydrique est représentée en haut, à droite, par les symboles verts ; cette relation est étroite et presque linéaire. Ces mesures ont été réalisées en Guyane (site de Paracou) sur un jeune arbre de l’espèce Simarouba amara. Dans cette expérimentation, la transpiration a été mesurée au moyen d’un capteur de flux de sève brute inséré dans le tronc de l’arbre et le potentiel hydrique grâce à une chambre à pression.
Toutefois, ce modèle n’est pas complètement réaliste, car une plante possède différents organes, des bifurcations, des branches, rameaux et racines. Des modèles plus complexes sont alors construits, calqués sur cette architecture, comme celui qui est représenté sur la figure 4.
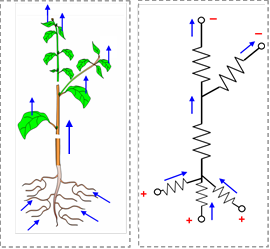
Figure 4. Un modèle de fonctionnement hydrique, plus réaliste que le précédent (figure 2), est calqué sur l’architecture de la plante, représentée à gauche. Ce modèle, représenté à droite, prend en compte les différents organes (racines, tige et feuilles) d’une plante et leurs ramifications. Les flèches indiquent les flux d’eau dans les différentes résistances hydrauliques, certaines étant connectées en série, d’autres en parallèle.
Ce type de modèle permet en particulier d’expliquer le fonctionnement hydrique du houppier d’un arbre. La figure 5 présente les variations de potentiel hydrique mesuré dans les feuilles (ici, ce sont des aiguilles) d’un jeune épicéa dans une forêt, en Lorraine. Ces mesures ont été réalisées à plusieurs niveaux de hauteur en utilisant des échafaudages dressés autour de l’arbre. On voit clairement que le potentiel hydrique, qui est maximal et stable pendant la nuit (c’est le potentiel de base), diminue dans la journée pour attendre les valeurs les plus basses entre 12 et 14 heures, donc au plus chaud de la journée, puis remonte progressivement ensuite. On constate aussi que le potentiel hydrique n’est pas identique à tous les niveaux de hauteur : il est le plus négatif dans les niveaux supérieurs de l’arbre, dans la partie ensoleillée du houppier, donc soumise à une forte demande climatique. Au contraire, les niveaux inférieurs, le plus souvent à l’ombre des arbres voisins, montrent des valeurs de potentiel moins négatives. En conséquence, les flux d’eau sont plus élevés dans les branches supérieures que les branches inférieures d’un arbre.
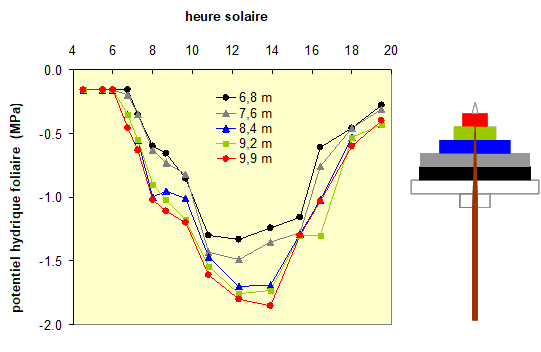
Figure 5. Variations du potentiel hydrique foliaire dans le houppier d’un jeune épicéa de 12 m de hauteur, mesuré à 5 niveaux de hauteur. Les couleurs des 5 courbes correspondent à celles des différents étages du houppier de l’arbre, schématisé à droite. Au début des mesures, pendant la nuit, le potentiel hydrique ne varie pas, il est environ égal à -0,2 MPa. Après le lever du soleil, vers 6 h du matin, les potentiels hydriques diminuent pour atteindre leur niveau le plus bas entre 12 et 14 h, les valeurs les plus faibles étant atteintes dans les parties supérieures de la couronne (niveaux rouge, vert et bleu), alors que les parties inférieures restent à des valeurs moins faibles (niveaux gris et noir). Les potentiels hydriques remontent ensuite progressivement au cours de la nuit suivante. Ces mesures ont été réalisées au moyen d’une chambre à pression. D’après Granier et Claustres (1989).
3.3. La plante en régime de flux non conservatif.
Nous avons vu que la transpiration consistait en un mouvement d’eau s’effectuant dans un circuit ouvert : l’eau qui entre par les racines fines finit par repartir, presque intégralement[5] vers l’atmosphère. Souvent, surtout lorsque la demande climatique est importante, sous un fort ensoleillement et des températures élevées, la transpiration des feuilles est supérieure à la quantité d’eau que peuvent absorber les racines. Dans de telles conditions, la plante se dessèche partiellement dans la journée. Mais en général, à partir de la fin d’après-midi jusqu’à la nuit, l’absorption racinaire devient supérieure à la transpiration et la plante récupère l’eau perdue, ses organes desséchés peuvent se réhydrater. Ainsi, tant en début qu’en fin de journée, la plante fonctionne en régime de flux non conservatif, comme cela est schématisé sur la figure 1. Certains tissus de certains organes chez la plupart des plantes ont en effet la capacité de se déshydrater et se réhydrater en fonction des variations de leur potentiel hydrique. Lorsque ces variations de teneur en eau concernent des tissus élastiques, il est assez facile de mesurer ce phénomène, car leur dimension varie avec celles de leur teneur en eau. Un exemple en est présenté sur la figure 6. Dans cette expérimentation, des capteurs de mesure automatiques ont enregistré les variations de diamètre sur un tronc et sur une branche d’un hêtre en forêt. On constate que dans la journée ces organes se contractent à partir de 8 heures jusque vers 14 heures, puis se dilatent ensuite aux heures moins chaudes jusqu’au milieu de la nuit qui suit. On constate un retard d’une heure à une heure et demie sur le rayonnement solaire des variations de diamètre des deux organes mesurés. De telles variations de diamètre, conséquence des variations de teneur en eau, sont observables aussi bien sur les tiges, les branches, les feuilles et les racines. Ces tissus échangeurs d’eau sont appelés des réservoirs internes. Ce phénomène est observable aussi sur des fruits[6].
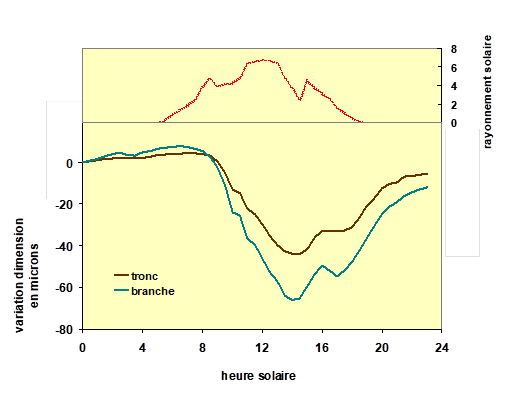
Figure 6. Mesures automatiques des variations de diamètre d’un tronc et d’une branche sur un jeune hêtre dans une forêt, lors d’une journée estivale. On remarque que ces deux organes se rétractent simultanément dans la journée, jusqu’à environ 14 heures, puis retrouvent progressivement dans la nuit leurs dimensions initiales. On note que la branche se rétracte un peu plus que le tronc. On a aussi représenté sur cette figure (en haut, courbe en rouge) les variations de rayonnement solaire, moteur de la transpiration de l’arbre, mesuré à proximité. Les variations de dimension de ces organes de l’arbre montrent un retard d’environ 1,5 à 2 heures sur celles du rayonnement. Mesures réalisées par Hervé Cochard, INRA.
Ces observations ont amené les chercheurs à introduire, dans le modèle électrique représenté sur la figure 4, différents compartiments (les réservoirs internes) pouvant se vider ou se remplir, en y incorporant une ou plusieurs capacitances. En effet, en électricité, une capacitance peut stocker ou libérer du courant électrique selon la variation de différence de potentiel électrique (U) à ses bornes : la capacitance se décharge lorsque U augmente, et inversement. La figure 7 représente un tel modèle appliqué à un des organes de la plante.
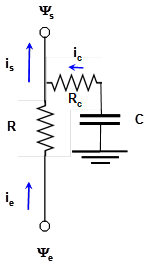
Figure 7. Modèle de fonctionnement hydraulique prenant en compte les réservoirs d’eau échangeable dans la plante. Ici un seul organe est représenté, par exemple la tige ou le tronc, de résistance R. L’eau circule dans l’organe au travers de sa résistance R, sous l’effet de la différence de potentiel hydrique (Ys - Ye). Si Ys – Ye augmente, la capacitance C se décharge (l’organe végétal considéré perd de l’eau) et libère un flux d’intensité ic qui s’ajoute au flux ie qui entre dans l’organe, pour donner is, le flux qui en sort.
Un stress majeur pour la plante : la sécheresse du sol
Que se passe-t-il lorsque le sol se dessèche trop fortement ? L’eau se trouve de plus en plus fortement liée aux particules de sol, comme dans une éponge très peu humide, qu’il faut presser fortement pour en extraire l’eau. Si, dans un sol humide, la plante peut sans trop de difficultés absorber l’eau car il y a peu de freins à sa circulation, progressivement, au bout de quelques jours à quelques semaines sans pluie ou sans arrosage, un seuil de contrainte hydrique est atteint. La figure 8 montre un exemple de cette chronologie sur un jeune arbre : le seuil de sécheresse, lié à la diminution du potentiel hydrique, déclenche un signal interne qui ferme les stomates de ses feuilles, limitant ainsi la transpiration, mais aussi réduisant d’autres fonctions vitales, comme la photosynthèse et la croissance. Cette fermeture des stomates est seulement partielle au début d’une période de sécheresse et peut devenir totale au plus fort du dessèchement. Dans un même temps, le potentiel hydrique diminue, aussi bien le potentiel de base (la nuit) que le potentiel hydrique minimum en milieu de journée. Dans ces conditions, les organes de la plante vont se dessécher un peu plus chaque jour. La nuit, la plante va pouvoir se réhydrater, mais seulement partiellement. En effet comme il a été vu plus haut, la plante ne pourra au mieux au cours de la nuit que retrouver le potentiel hydrique du sol.
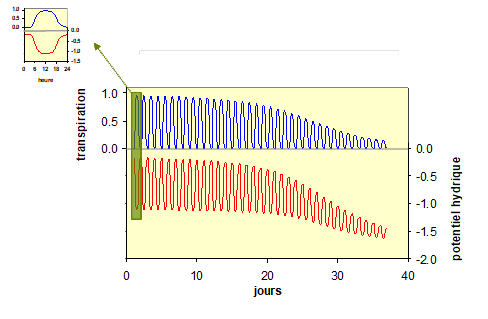
Figure 8. Variations modélisées de la transpiration (courbe en bleu) et du potentiel hydrique (en rouge) d’un arbre lors de 36 journées ensoleillées successives, en partant d’un sol humide. Chaque oscillation correspond à une journée : la nuit la transpiration est nulle et le potentiel hydrique peu négatif. Au milieu de la journée la transpiration est maximale et le potentiel hydrique est à son minimum. Voir le détail de la première journée, dans le petit cadre en haut à gauche. En l’absence de pluie, l’eau du sol est progressivement consommée par l’arbre. Au bout de 15 jours environ, l’eau du sol devient difficile à extraire : le potentiel hydrique de l’arbre diminue, aussi bien de jour comme de nuit, mais les mécanismes de protection l’arbre réduisent progressivement sa transpiration par la fermeture fermant de ses stomates.
Enfin, la diminution du potentiel hydrique causée par la sécheresse (mais aussi par d’autres stress comme le gel ou des attaques d’insectes) pourra avoir aussi pour conséquence la rupture des colonnes de sève brute dans les trachéïdes et les vaisseaux du xylème, comme un fil qui se rompt lorsque la traction entre ses deux extrémités devient trop forte : c’est la cavitation. Cette rupture des petites colonnes d’eau dans les éléments conducteurs est très rapidement suivie par le remplissage du vide créé avec de la vapeur d’eau : c’est l’embolie. L’embolie rend la circulation de l’eau impossible, tout comme elle bloque la circulation dans un petit vaisseau sanguin de notre corps. Cet état peut provoquer le dessèchement irréversible des feuilles, ainsi que parfois celui des petits rameaux des arbres, voire de branches entières. Feuilles et rameaux peuvent alors se détacher de la plante. La plupart du temps, l’embolie n’est pas réversible et représente un affaiblissement de la plante, parfois létal (voir Chap. II.7).
Sommaire :
Dans le chapitre :


