Efficacité d’utilisation de l’eau par les plantes en fonction de leur métabolisme photosynthétique
Introduction
La consommation en eau des cultures est étroitement liée à leur production de biomasse car le dégagement de vapeur d’eau et l’absorption du CO2 sont liés à la porosité de l’épiderme des feuilles, que l’on quantifie par la mesure de la conductance stomatique (voir Chap. II.5). Il existe toutefois des différences entre les espèces végétales, qui peuvent être classées en trois types métaboliques : les plantes en C3, en C4 et les CAM (voir photo). Ces types se différencient par la manière dont le CO2 de l’air est fixé par photosynthèse après son absorption et sa dissolution en phase aqueuse, où il coexiste sous trois formes : comme CO2 dissous, comme bicarbonate HCO3- et comme carbonate CO3--.

Photo : Des cultures de type C3, C4 et CAM. A gauche, le colza (Brassica napus), une plante en C3, au centre, le maïs (Zea mays), une plante en C4 et à droite, l’ananas (Ananas comosus), une plante de type CAM.
Les trois types métaboliques
Les espèces en C3 constituent la grande majorité des espèces. Les principales espèces cultivées (blé, riz, orge, soja, colza) sont en C3 (à l’exception de certaines espèces d’origine tropicale comme le maïs ou le sorgho) ainsi que toutes les légumineuses et tous les arbres. Chez les espèces en C3, le carbone du CO2 dissous est fixé sur un sucre en C5, le Ribulose bis Phosphate (RubP) pour former deux molécules à trois atomes de carbone (C3) d’acide phosphoglycérique PGA :
![]() RubP + CO2 2 PGA (1)
RubP + CO2 2 PGA (1)
Le RubP réagit aussi avec l’oxygène de l’air pour donner une molécule en C3 (PGA) et une autre en C2, le phosphoglycolate (P-glycolate) qui ultérieurement libère une molécule de CO2. C’est donc une forme de respiration apparente (différente de la respiration mitochondriale[1]), et comme la régénération du RubP nécessite de la lumière, elle est appelée photorespiration :
![]() RubP + O2 PGA + P-glycolate (2)
RubP + O2 PGA + P-glycolate (2)
Les réactions (1) et (2) sont catalysées par la même enzyme, appelée pour cette raison RubP carboxylase-oxygénase, en abrégé Rubisco. Le rapport vC/vO (de la vitesse de carboxylation à celle d’oxygénation) est proportionnel au rapport de concentrations [CO2] / [O2] en phase dissoute au niveau de la Rubisco (dans le chloroplaste) et diminue quand la température augmente. Le résultat est que dans l’atmosphère actuelle à température de 25 °C, environ 1/3 du carbone fixé par photosynthèse est perdu par photorespiration. Le cycle métabolique du P-glycolate photorespiratoire permet de recycler les ¾ du carbone du CO2 émis après oxygénation du RubP, ce qui a un coût énergétique non négligeable.
Les espèces en C4 qui sont d’apparition tardive (7 à 10 millions d’années) sont représentées par des espèces cultivées d’origine tropicale comme le maïs, le sorgho, le mil, le millet, la canne à sucre (beaucoup de graminées tropicales sont en C4, mais pas le riz qui est en C3). Ces espèces se caractérisent par deux carboxylations successives dans deux types de cellules différents. Dans le cytoplasme des cellules du mésophylle (milieu de la feuille), le carbone du bicarbonate HCO3- est fixé sur le phospho-énol-pyruvate (PEP), une molécule en C3, pour donner un acide organique en C4, l’acide oxalo-acétique (OAA) et du phosphate inorganique Pi :
![]() PEP + HCO3- OAA + Pi (3)
PEP + HCO3- OAA + Pi (3)
Cette réaction est catalysée par le PEP carboxylase, une enzyme à forte affinité pour le HCO3- et sans affinité pour l’oxygène. L’OAA, réduit en malate[2], migre ensuite vers les cellules de la gaine entourant les vaisseaux de la feuille et libère, après décarboxylation, du CO2 qui s’accumule puis diffuse vers les chloroplastes de la gaine pour être fixé sur le RubP par la Rubisco par une deuxième carboxylation de type (1) comme chez les plantes en C3. L’avantage de ce système de double carboxylation est de concentrer le CO2 dans les cellules de la gaine, augmentant le rapport CO2/O2 d’un ordre de grandeur, ce qui supprime quasiment la photorespiration et augmente d’autant la fixation nette de carbone. L’inconvénient est un coût énergétique supérieur pour permettre la régénération du RubP qui nécessite une molécule nommée adénosine triphosphate, en abrégé ATP[3] fournie par la phase photochimique de la photosynthèse, ce qui peut être un facteur limitant dans des conditions de faible éclairement. La structure anatomique des feuilles de plantes C4 présente deux manchons cellulaires (mésenchyme chlorophyllien et gaine périvasculaire, voir Fig. 1), ce qui est très favorable à la retenue du CO2 près des sites catalytiques de la Rubisco.
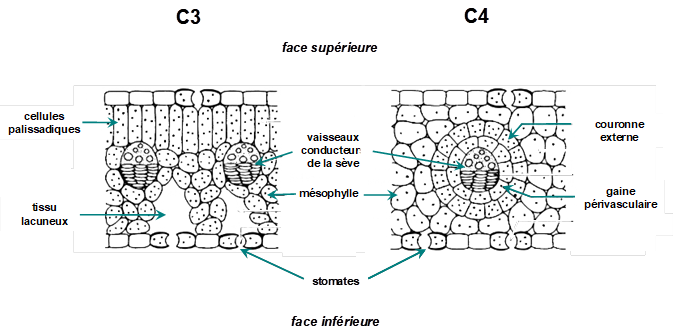
Figure 1 : coupe transversale de feuilles de plantes en C3 et en C4
Les espèces de type CAM (abréviation de Crassulacean Acid Metabolism ; en français : métabolisme acide des crassulacées) sont des espèces de plantes succulentes qu’on rencontre chez les crassulacées, les cactacées, les euphorbiacées et de nombreuses autres familles, dont le métabolisme est une variante du type C4. Elles présentent la particularité d’ouvrir leurs stomates la nuit et de les fermer pendant la plus grande partie du jour, ce qui limite les pertes d’eau par transpiration. La première carboxylation se produit la nuit par la réaction (3), l’OAA est réduit en acide malique qui s’accumule dans les grandes vacuoles de ces plantes. Le jour, l’acide malique après décarboxylation libère du CO2 qui est repris par la deuxième carboxylation (1) commune à toutes les espèces. Ainsi chez les CAM les deux carboxylations (par PEPcase et Rubisco) sont séparées dans le temps alors que chez les C4 elles sont séparées dans l’espace. Les agaves, Aloes, euphorbes, Sedum sont en CAM ainsi que quelques espèces cultivées comme l’ananas, le sisal ou le Kalanchoe, une plante ornementale.
L’avantage des CAM est de limiter leurs pertes en eau par la fermeture stomatique de jour, leur inconvénient est d’avoir une capacité de fixation du carbone limitée par le stockage nocturne d’acides dans les vacuoles, et aussi comme chez les C4 une dépense supplémentaire d’énergie pour régénérer le RubP. De plus, la plupart des espèces CAM sont dites facultatives, en ce sens qu’elles peuvent fonctionner comme des C3 lorsque le sol est bien humide, en ouvrant leurs stomates de jour. Ces limitations peuvent être en partie levées dans des cultures d’espèces CAM à haute densité, leur permettant d’atteindre des indices foliaires de 2 à 4 (Nobel, 1991). Nobel a ainsi montré qu’on pouvait atteindre 26 tMS[4] ha-1 an-1 pour 5 espèces CAM cultivées (avec une précipitation annuelle de 880 mm), à rapprocher de 35 tMS ha-1 an-1 pour les meilleures cultures en C3 et de 49 tMS ha-1 an-1 pour les meilleures en C4.
Différences d’efficacité d’utilisation de l’eau (EUE) entre les types métaboliques
Quelles conséquences ont ces types métaboliques sur la quantité d’eau nécessaire pour produire de la biomasse ? On peut exprimer l’assimilation de CO2 sous la forme :
![]() (4)
(4)
Où ![]() est la conductance stomatique (porosité de l’épiderme) pour le CO2, Ca et Ci les concentrations en CO2 de l’air externe et interne à la feuille. Si Ca et Ci sont exprimées en parties par million en volume (ppm) et
est la conductance stomatique (porosité de l’épiderme) pour le CO2, Ca et Ci les concentrations en CO2 de l’air externe et interne à la feuille. Si Ca et Ci sont exprimées en parties par million en volume (ppm) et ![]() en mol m-2 s-1, alors A est en µmol m-2 s-1. La perte d’eau par transpiration peut s’exprimer de la même façon comme la sortie de vapeur d’eau de la feuille après évaporation :
en mol m-2 s-1, alors A est en µmol m-2 s-1. La perte d’eau par transpiration peut s’exprimer de la même façon comme la sortie de vapeur d’eau de la feuille après évaporation :
![]() (5)
(5)
Où ![]() est la conductance stomatique pour la vapeur d’eau, ei et ea les pressions partielles de vapeur d’eau de l’air interne et externe à la feuille, p la pression atmosphérique. Si ei et ea sont exprimées en kPa, p en MPa, et
est la conductance stomatique pour la vapeur d’eau, ei et ea les pressions partielles de vapeur d’eau de l’air interne et externe à la feuille, p la pression atmosphérique. Si ei et ea sont exprimées en kPa, p en MPa, et ![]() en mol m-2 s-1, E est en mmol m-2 s-1. L’air interne à la feuille est pratiquement saturé en vapeur d’eau à cause de la forte hydratation des cellules et on a donc ei ~ es(Tf) qui est la pression de vapeur saturante à la température de la feuille.
en mol m-2 s-1, E est en mmol m-2 s-1. L’air interne à la feuille est pratiquement saturé en vapeur d’eau à cause de la forte hydratation des cellules et on a donc ei ~ es(Tf) qui est la pression de vapeur saturante à la température de la feuille.
En faisant le rapport des équations (4) et (5) on obtient le rapport entre le CO2 fixé et l’eau consommée par la feuille, ou efficacité d’utilisation de l’eau au niveau foliaire :
![]() (6)
(6)
Le rapport ![]() est une constante égale au rapport des coefficients de diffusion du CO2 et de la vapeur d’eau dans l’air, qui sont inversement proportionnels à la racine carrée de leur masse moléculaire (cf. les gaz légers diffusent plus vite), par suite :
est une constante égale au rapport des coefficients de diffusion du CO2 et de la vapeur d’eau dans l’air, qui sont inversement proportionnels à la racine carrée de leur masse moléculaire (cf. les gaz légers diffusent plus vite), par suite : ![]() . La pression de vapeur d'eau interne à la feuille ei augmente avec sa température, qui est fonction du rayonnement absorbé, de l’ouverture stomatique et des conditions atmosphériques, mais est souvent proche de la température de l’air lorsque la feuille est bien hydratée. Le principal facteur de variation de l’EUE est donc Ca – Ci, souvent mis sous la forme Ca(1-Ci/Ca) pour faire apparaître le rapport des concentrations de CO2 à l’intérieur de la feuille et à l’extérieur.
. La pression de vapeur d'eau interne à la feuille ei augmente avec sa température, qui est fonction du rayonnement absorbé, de l’ouverture stomatique et des conditions atmosphériques, mais est souvent proche de la température de l’air lorsque la feuille est bien hydratée. Le principal facteur de variation de l’EUE est donc Ca – Ci, souvent mis sous la forme Ca(1-Ci/Ca) pour faire apparaître le rapport des concentrations de CO2 à l’intérieur de la feuille et à l’extérieur.
La forte affinité de la PEP carboxylase des espèces en C4 leur permet d’abaisser la concentration interne Ci beaucoup plus que chez les plantes en C3, il en résulte que Ci/Ca vaut environ 1/3 chez les plantes C4 contre 2/3 chez les plantes C3, faisant doubler (1-Ci/Ca) entre les C3 et les C4.
Ainsi, toutes choses égales par ailleurs, une espèce en C4 fixe plus de carbone qu’une espèce en C3 pour une même consommation en eau. Au champ, la différence est souvent moindre que celle calculée ici, pour deux raisons. D’une part quand on passe de la feuille au couvert l’évaporation du sol entraîne des pertes en eau supplémentaires, et la forte surface foliaire de la culture tend à gommer les différences. D’autre part, le maximum de surface foliaire est atteint pour le blé au printemps dans des conditions de demande évaporative modérée, et en plein été pour le maïs, lorsque la demande évaporative est forte.
On cite souvent le chiffre de 500 L d’eau nécessaires pour produire un kg de grains, soit une EUE de 2 g/kg. Dans des unités agronomiques le rendement est exprimé en kg/ha et la consommation en eau en mm ou L/m2 ou kg/m2 ; comme 1 g/m2 = 10 kg/ha et 1 kg/m2 = 1 mm, 2 g/kg = 20 kg ha-1 mm-1. C’est la valeur maximale citée par Passioura (2006) pour du blé en Australie du sud. Les valeurs d’EUE sont évidemment plus élevées quand on se réfère à la biomasse aérienne plutôt qu’aux grains. Ainsi Neal et al. (2005) trouve près de Sydney des valeurs d’EUE allant de 13,5 à 43 kg ha-1 mm-1 pour des fourrages, la valeur la plus faible étant pour une légumineuse du genre Vigna et la plus élevée pour du maïs. En Europe près de Berlin, Mueller et al. (2005) cite des valeurs d’EUE pour la biomasse aérienne supérieures à 40 kg ha-1 mm-1 pour le maïs, d’environ 30 pour le blé et de 15 à 28 pour diverses cultures fourragères. Récemment Beer et al. (2009) ont utilisé les mesures de flux de CO2 et de vapeur d’eau pour évaluer l’EUE au dessus de divers couverts végétaux. Ils ont trouvé pour les cultures et les prairies une moyenne de 2,9 gC/kg(eau), rapport entre la quantité de carbone fixée par photosynthèse et l’eau évapotranspirée. Si l’on prend un indice de récolte[5] de 0,4 et un rapport carbone sur matière sèche de 0,45, ce chiffre correspond à 26 kg (grains) ha-1 mm-1 soit une valeur un peu supérieure à celle indiquée par Passioura. Il est normal de trouver une EUE supérieure en climat plus frais : en effet dans l’équation (6) figure au dénominateur (ei – ea), qui augmente sous climat chaud et sec, ce qui a incité Beer et al. (2009) à multiplier EUE par le déficit en vapeur d’eau de l’air ![]() qui est une bonne approximation de (ei – ea). Ce procédé permet de calculer une EUE dite intrinsèque, dépendant seulement de la photosynthèse et de l’ouverture stomatique et pas du climat. On trouvera dans le tableau ci-dessous un résumé de données sur feuilles, plantes entières et couverts végétaux.
qui est une bonne approximation de (ei – ea). Ce procédé permet de calculer une EUE dite intrinsèque, dépendant seulement de la photosynthèse et de l’ouverture stomatique et pas du climat. On trouvera dans le tableau ci-dessous un résumé de données sur feuilles, plantes entières et couverts végétaux.
Tableau. Efficacités d’utilisation de l’eau chez divers types métaboliques. « Théorique » se réfère au calcul de l’équation (6) en supposant ei – ea = 19 hPa (température de 25 °C et humidité relative de l’air de 50%), Ca = 380 ppm, Ci/Ca = 2/3 pour les C3 et 1/3 pour les C4. Les chiffres mesurés sur plantes en pot correspondent à la production de biomasse, qui inclut les pertes par respiration nocturne, alors que les mesures sur feuille sont fondées sur les échanges de CO2 et de vapeur d’eau. Toutes les données ont été converties dans la même unité.
|
type de |
théorique |
mesuré |
plante |
culture |
|
C3 |
6 |
4 |
1-2 |
1,4 - 3 |
|
C4 |
12 |
8 |
3-4 |
3 - 4 |
|
CAM |
|
|
8-20 |
6 |
|
unité |
gMS / kgeau |
gMS / kgeau |
gMS / kgeau |
gMS / kgeau |
Conclusion
Quelles sont les perspectives d’améliorer la production des plantes cultivées lorsque l’eau devient rare? On peut d’abord augmenter l’eau disponible pour la plante, en ajustant la date de semis à la saison des pluies, en améliorant le système racinaire pour mieux exploiter la réserve en eau du sol, ou en diminuant l’évaporation du sol (voir Chap. III.1 et IV.1). On peut ensuite augmenter l’efficacité d’utilisation de l’eau (EUE) en diminuant la conductance stomatique ou en améliorant la fixation de CO2 (ce qui entraîne une diminution de Ci dans l’équation (6)). On peut noter au passage que l’augmentation de teneur en CO2 de l’air entraîne une légère fermeture stomatique et une augmentation de photosynthèse qui augmente l'EUE ; tout n'est donc pas négatif dans le changement global. Enfin on peut aussi augmenter par sélection génétique l’indice de récolte, le rapport entre la biomasse récoltée et la biomasse totale de la plante.
Passioura (2006) a fourni une analyse réaliste de ce qui peut être espéré dans les prochaines années. Croire qu’on peut faire pousser une plante avec très peu d’eau en modifiant quelques-uns de ses gènes relève du fantasme et doit être dénoncé. La génomique fonctionnelle peut apporter beaucoup dans l’avenir pour améliorer le rendement en conditions hydriques limitantes, mais seulement dans un contexte bien compris sur les plans biochimique, physiologique et agronomique (voir Chap. III.1). Les perspectives d’amélioration concernant la feuille et la photosynthèse sont modérées (on espère un jour transformer le riz en espèce en C4 mais il s’agit d’un projet à long terme : http://irri.org/c4rice, Kajala et al., 2011). Dans l'immédiat, on peut jouer sur la dynamique d’établissement de la surface foliaire, sur la profondeur d’enracinement, sur la phase reproductive critique pour le rendement. L’objectif est d’optimiser l’utilisation de la réserve en eau du sol de façon à laisser assez d’eau à la floraison pour permettre la germination du pollen, la fécondation et le développement du grain. Bien sûr si l’irrigation est possible, un complément peut être apporté à ce moment. On peut aussi ralentir la phase de sénescence (gènes permettant de « rester vert ») mais ce sera bénéfique seulement si la culture dispose d’assez d’eau pour pouvoir en profiter. On voit alors que le paysan qui dépend seulement de la pluie devra adapter sa densité de semis et de manière générale le développement de la surface foliaire de sa culture en fonction de la quantité d’eau espérée (voir Chap. III.1 et III.2). Il est préférable d’avoir une récolte minimale en année sèche plutôt que pas de récolte du tout, mais le prix à payer est un rendement moindre en année humide. Les paysans tunisiens ont bien compris ce principe en ajustant la densité de leurs oliviers aux pluies disponibles, qui diminuent du nord au sud du pays (voir Chap. IV.1).
Références bibliographiques
Beer C. et al. (2009) Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Global Biogeochemical Cycles, 23, GB2018, doi:10.1029/2008GB003233.
Kajala K et al. (2011) Strategies for engineering a two-celled C4 photosynthetic pathway into rice. Journal of Experimental Botany Advance Access published Feb. 18, 2011, 1-10.
Mueller et al. (2005) Above ground biomass and water use efficiency of crops at shallow water tables in a temperate climate. Agricultural Water Management, 75, 117–136.
Neal JS, Fulkerson WJ, Hacker RB (2011) Differences in water use efficiency among annual forages used by the dairy industry under optimum and deficit irrigation. Agricultural Water Management, 98, 759–774.
Nobel P.S. (1991) Achievable productivities of certain CAM plants: basis for high values compared with C3 and C4 plants. New Phytologist, 119: 183–205.
Passioura JR (2006) Increased crop productivity when water is scarce- from breeding to field management. Agricultural Water Management, 80, 176-196.
[1] La respiration mitochondriale a son siège dans les mitochondries de cellules. Commune à tous les organismes vivants, elle leur fournit l’énergie dont ils ont besoin pour assurer leurs fonctions vitales, dont la croissance.
[2] Le malate est un sel de l’acide malique
[3] l’ATP permet à une cellule d’emmagasiner de l’énergie ou de la libérer très rapidement
[4] tMS : tonnes de matière sèche
[5] L’indice de récolte est le rapport entre le rendement commercialisable d'une culture, par exemple les grains et la quantité totale de biomasse produite par la culture
Sommaire :
Dans le chapitre :


