Autres indicateurs de l’état hydrique des plantes
La déshydratation des organes de la plante et la chute de son potentiel hydrique par rapport à son état optimal entraînent des modifications physiologiques et morphologiques, parfois visibles à l’œil nu. En dehors de la déshydratation des organes de la plante, qui provoque souvent leur contraction parallèlement à la diminution du potentiel hydrique, d’autres effets peuvent être mis en évidence par des méthodes variées, dont nous donnerons brièvement ci-dessous une liste non exhaustive. Certains travaux ont porté sur la comparaison de la sensibilité et de la fiabilité de différentes méthodes de mesure de l’état hydrique des plantes, par exemple l’article de Katerji et al. (1988) sur la tomate et celui de Dwyer et Stewart (1984) sur le maïs, la finalité de ces travaux étant la recherche d’un critère de déclenchement de l’irrigation de cultures. Il ressort de diverses études que la mesure du potentiel de base est le critère le plus fiable, mais il a pour inconvénient la contrainte forte des conditions de mesure, réalisées en fin de nuit.
5.1 Critères visuels
Les variations de l’état hydrique des organes d’une plante entraînent souvent des changements de leur apparence, par exemple dans le port des tiges qui s’affaissent plus ou moins ou dans l’aspect extérieur de la forme des feuilles. Par exemple, chez le maïs et le riz, il existe une assez bonne corrélation entre l’enroulement des feuilles et leur potentiel hydrique : l’agriculteur peut ainsi avec un peu d’exercice déterminer «à l’œil» une valeur assez précise du potentiel hydrique foliaire qui permettra de décider ou non d’irriguer.
5.2. La conductance stomatique et la photosynthèse
La sécheresse induit chez de nombreuses espèces [5] la fermeture des stomates (voir chapitre II.5), qui réduit la transpiration de la plante ainsi que sa photosynthèse et en cascade entraîne d’autres conséquences physiologiques. Il existe des appareils, appelés les poromètres, permettant de mesurer la conductance stomatique, tant au laboratoire que sur le terrain. Ces équipements sont plutôt réservés au domaine de la recherche. Le poromètre, dont nous ne décrivons pas le principe ici, est une petite chambre de mesure dans laquelle est enfermée une feuille ou seulement une portion du limbe de celle-ci. Cet appareil, qui contient divers capteurs (température, rayonnement, humidité) permet de mesurer les variations de conductance stomatique au cours d’une journée sur différentes plantes d’une population, et de détecter le moment où la régulation stomatique se met en place.
5.3. La température des feuilles
La fermeture des stomates limite, voire bloque, la transpiration des feuilles. En considérant le bilan d’énergie d’une feuille, on peut montrer (voir chapitre II.5) que si, ni le rayonnement solaire, ni l’état de la masse d’air (température, humidité, vent) ne varient, la température des feuilles augmente et cette fermeture des stomates provoque une augmentation de température qui devient devient supérieure à celle de l’air. Cet écart de température est modeste, de l’ordre de un à quelques °C, mais mesurable avec des équipements relativement simples (radiothermomètres à infrarouge) dont la précision et la justesse s’améliorent rapidement. On trouve maintenant dans le commerce des radiothermomètres, souvent utilisés pour effectuer des mesures de température dans les habitations afin d’établir des diagnostics thermiques. On mesure avec ceux-ci la température des murs, des sols et des plafonds, celle des radiateurs de chauffage etc. Ils peuvent aussi être utilisés dans des parcelles agricoles, des vergers ou des forêts. La Fig. 15 montre un exemple de mesures réalisées sur des oliviers en climat chaud et sec, soumis à différents traitements d’irrigation. On y voit que les feuilles des arbres bien irrigués sont à une température d’environ 2°C plus faible que celle des arbres du traitement le plus sec. Certains satellites d’observation de la terre en sont équipés (Landsat-5 par exemple) permettant de détecter avec une bonne résolution spatiale (de l’ordre de quelques mètres) les zones soumises à la sécheresse.
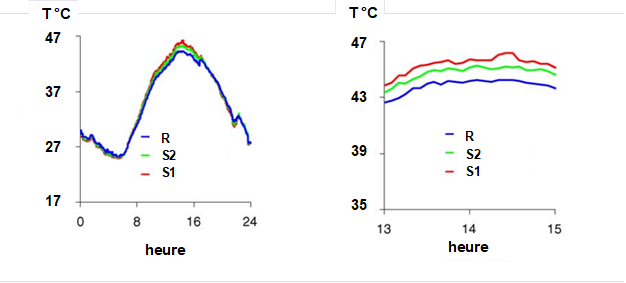
Figure 15. Température du feuillage d’oliviers en plantation dans un verger au sud de l’Espagne recevant différentes quantités d’eau d’irrigation. R, les arbres reçoivent 2,8 mm/jour, 1,2 mm/jour pour S2 et 0,7 mm/jour pour S1. A gauche, une journée complète (le 25 juillet). A droite, le détail des températures aux heures les plus chaudes. Les températures sont mesurées au moyen de capteurs à infrarouge installés de façon fixe au-dessus des arbres sur des potences. Repris de Sepulcre-Cantó (2006).
La différence de température entre des couverts végétaux de même nature en fonction de leur état hydrique a conduit à la construction d’un indice susceptible d’être utilisé pour le pilotage de l’irrigation. Cet indice est basé sur la mesure de la température des couverts végétaux par thermométrie infrarouge, réalisée parfois par avion et le plus souvent par des satellites dédiés. On part de l’idée suivante : un couvert végétal est à a une température T, qui est comprise entre deux bornes : une valeur maximale Tmax, qui serait celle d’un couvert de mêmes caractéristiques physiques mais totalement sec et une valeur minimale Tmin, celle d’un couvert identique en total confort hydrique.
On définit alors l’index CWSI (Crop water Stress Index) :
CWSI = (T-Tmin) / (Tmax – Tmin)
On a dès lors deux problèmes à résoudre :
- calculer CWSI : pour cela on remplace T et Tmin par les valeurs de températures radiatives mesurées à la fois sur un couvert en cours de dessèchement et un couvert bien irrigué. On calcule Tmax à partir de l’hypothèse que le flux de chaleur sensible égale, dans ce cas, le rayonnement net mesuré (voir Chap. II.5).
- relier le CWSI aux mesures d’évapotranspiration relative comme dans le cas du potentiel de base. Si l’on disposait de mesures parfaites de T, Tmin et Tmax sur une vraie surface, la relation théorique serait : CWSI = 1-rET ou rET = 1 – CWSI.
La Fig. 16, d’après Itier et al. (1993), donne un exemple de la relation obtenue avec des mesures de terrain.
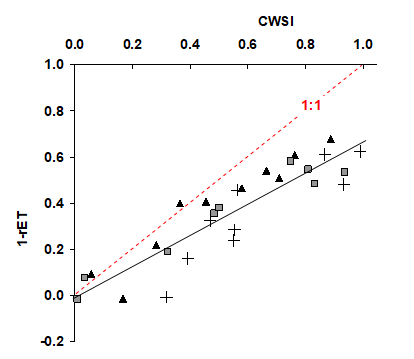
Figure 16. Relation entre l’indice CWSI et évapotranspiration relative (rET) sur une culture de soja en 1989 à Mas d’Asport (30-France) pour trois cycles de dessèchement.
Bien que les relations obtenues soient prometteuses, on peut observer sur cette figure que :
- d’une part il y a une forte dispersion des points autour de la relation statistique (trait noir). En effet, la mesure radiométrique pose des problèmes, comme par exemple le choix de l’incidence de visée pour la mesure, sachant qu’il faut éviter l’influence parasite de la température du sol sous le couvert végétal. Par ailleurs, la température maximale calculée n’est pas la température radiométrique des feuilles supérieures, vues par le capteur ;
- d’autre part la relation statistique trouvée s’écarte notablement de la pente 1:1.
On est encore loin de l’opérationnalité de ce type d’index développés par les promoteurs de la télédétection, rapprochée ou non.


