Expressions de l’état hydrique des végétaux
Plusieurs variables permettent de caractériser l’état hydrique de la plante ; nous en explicitons ci-dessous les plus importantes. Nous verrons aussi que certaines de ces grandeurs sont liées entre elles, parfois étroitement.
2.1 Grandeurs massiques
Ces grandeurs sont basées sur la mesure de la quantité d’eau contenue dans un échantillon. On en distingue principalement quatre : l’humidité pondérale, la teneur en eau, la teneur en eau relative et le déficit hydrique. Pour les grandeurs utilisant le poids sec d’un organe ou d’une plante comme référence, on suppose que celui-ci ne change pas lorsque l’on s’intéresse aux variations à court terme du contenu en eau.
Humidité pondérale
L’humidité pondérale (Hp) est le rapport entre le poids d’eau d’un échantillon (Peau) et son poids sec (Psec) :
Hp = Peau/ Psec (1)
Par exemple, si Hp est égal à 3, cela signifie que pour 1 g de matière sèche, l’échantillon contient 3 g d’eau. Psec est déterminé par passage à l’étuve à 70-80 °C (NB : des températures plus élevées risquent d’évaporer des composés organiques volatils) jusqu’à ce que ce poids devienne constant. La durée dépend de la nature et du volume des échantillons : de 24 heures pour des feuilles à quelques jours pour des échantillons de bois volumineux. Hp peut aussi être exprimé en pourcentage.
Teneur en eau
La teneur en eau (Te) est le rapport entre le poids d’eau d’un échantillon (Peau) et son poids frais au moment du prélèvement (Pfrais), exprimé en pourcentage :
Te = 100.(Peau/Pfrais) (2)
Cette relation peut aussi s’écrire :
Te = 100 . Hp / (1 + Hp)
Noter que par définition Pfrais = Peau + Psec. Noter aussi que la teneur en eau est une grandeur qui n’est pas très équilibrée, parce que l’on trouve le poids de l’eau à la fois au numérateur et au dénominateur ; cela « tamponne » les variations du contenu en eau, d’autant plus que Hp est grande.
Exemple : soit une feuille dont le poids sec est de 1 g. Si elle passe d’une Hp de 5 à 6 cela correspond à une augmentation de 20% de Hp ; Te, au contraire, passe de 5/6 = 0,83 à 6/7 = 0,86 soit une augmentation de 3% seulement.
Le tableau 1 présente un certain nombre de valeurs d’humidité pondérale et de teneur en eau mesurées sur différents organes de différentes espèces de plantes. Dans ce tableau, on peut voir que, mis à part les graines et le bois, les organes et tissus végétaux contiennent en général plusieurs fois leur poids sec en eau, parfois plus de 10 fois. Ces valeurs sont très variables, tant en fonction de l’organe (feuilles, tiges, racines peuvent avoir des Hp soit semblables, soit différentes) qu’avec l’espèce, l’âge de l’organe (habituellement, Hp diminue avec l’âge) et l’environnement dans lequel vit la plante. Par comparaison, l’humidité pondérale d’un sol, même humide, est en général bien inférieure aux valeurs de ce tableau, de l’ordre de 0,5 à 1.
Teneur en eau relative
La teneur en eau relative (TER), appelée aussi turgescence relative, est le rapport entre le poids d’eau contenu dans l’échantillon (Peau) au moment du prélèvement et le poids d’eau maximal (Pept) qu’il peut contenir, en conditions physiologiques «normales», quand il se trouve en équilibre avec de l’eau « libre » c’est-à-dire de l’eau pure à la même température (son potentiel hydrique est alors nul). Ce poids Pept correspond alors à un état hydrique maximal, qu’on appelle pleine turgescence. On a donc :
TER = 100 . (Peau/Pept) (3)
On voit donc que la TER ne peut être supérieure à 100%. En remplaçant Peau par sa valeur dans (1), on a aussi la relation :
TER = (Hp. Psec / Pept) (4)
Une même Hp correspond donc à différentes TER selon la valeur du poids sec et de la quantité d’eau à pleine turgescence.
Déficit hydrique
Le déficit hydrique (D, en %) se définit comme le complément à 100 de la TER :
D = 100 - TER (5)
Tout comme la TER, le déficit en eau ainsi défini, aura une valeur comprise entre 0 (ce sera la pleine turgescence) et 100 (son poids sera égal à son poids sec, il sera totalement sec).
La détermination de la TER implique qu’il faille, avant de le passer à l’étuve pour avoir son poids sec, porter l’échantillon à son état de pleine turgescence. Dans cet état, toutes les cellules de l’échantillon sont à leur degré maximal d’hydratation, en équilibre de potentiel hydrique avec de l’eau pure, à la même température et à la même pression. Les organes de loin les mieux adaptés à cette détermination sont les feuilles ou les disques foliaires. Avec une feuille, on peut atteindre un tel état en la plaçant dans une enceinte dont l’atmosphère est saturée en eau, le pétiole plongeant dans l’eau pure. Au lieu de prendre une feuille entière, on peut aussi prélever à l’emporte-pièce des disques foliaires que l’on fait réhydrater à partir d’une éponge saturée par exemple. Le petit complément en fin de § 3 indique quels phénomènes parasites peuvent venir compliquer cette détermination.
La mesure de la TER n’est pas toujours possible ; c’est le cas des fruits ou des cactées, qui ne peuvent être portés à la pleine turgescence, soit du fait de la lenteur du processus, soit du fait du pourrissement des tissus qui l’accompagne.
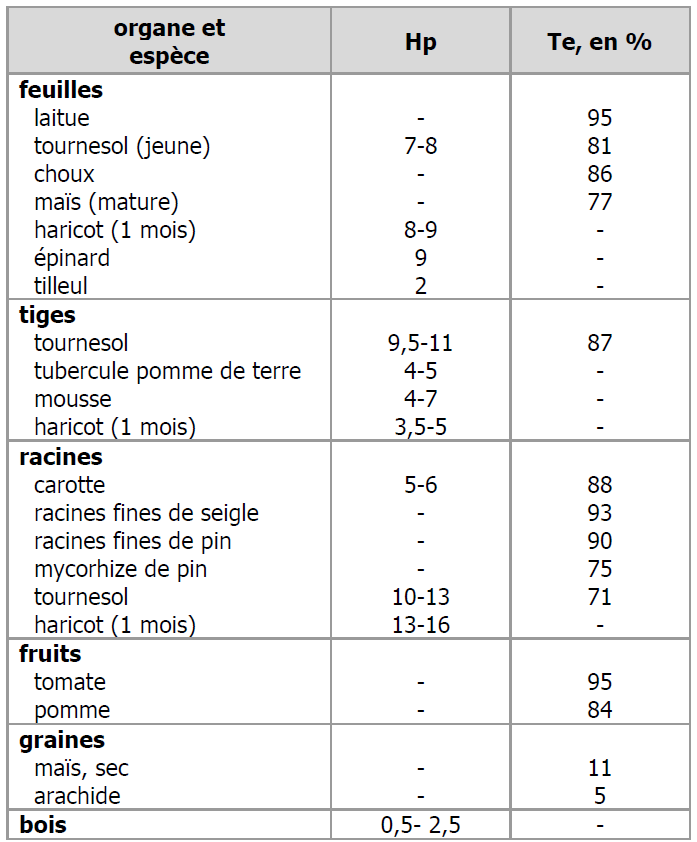
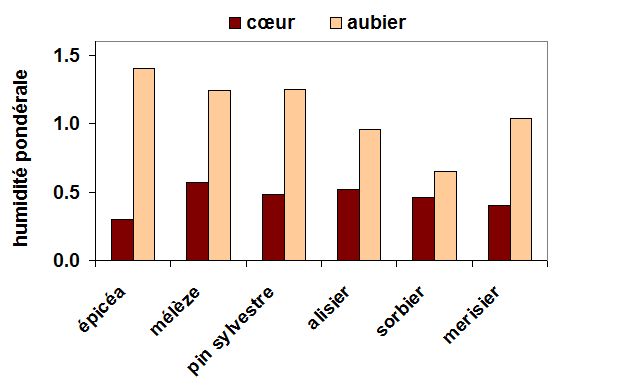
Figure 1. Humidité pondérale du bois d’aubier et du bois de cœur de troncs de quelques espèces forestières tempérées. D’après Polge (1964). Note : une valeur d'humidité pondérale égale à 1 signifie que ll'échantillon contient la même masse d'eau que de matière sèche.
Il existe chez certaines espèces, une forte hétérogénéité d’humidité au sein d’un même organe. C’est le cas en particulier du bois des arbres, lorsque qu’ils ne sont plus à leur stade juvénile. Le bois du tronc, mais aussi celui des grosses branches et des grosses racines, contient souvent une partie vivante (le bois d’aubier, qui est le tissu conducteur de la sève brute) et une partie morte (le bois de cœur, qui confère à l’arbre sa rigidité et sa résistance mécanique). Ces deux tissus montrent généralement de fortes différences de teneur en eau, ainsi que l’illustre la Fig. 1 pour différentes espèces d’arbres des forêts européennes tempérées. Il existe un contraste très marqué entre les deux tissus, le bois d’aubier étant beaucoup plus humide que le bois de cœur, celui-ci contenant parfois plus de 4 fois plus d’eau que ce dernier par unité de poids sec. Lorsque le bois de cœur se forme dans l’arbre, processus appelé duraminisation, il se charge en éléments minéraux et en molécules organiques inutiles voire néfastes au métabolisme de l’arbre et perd une partie de son eau, qui migre vers le bois d’aubier.
2.2 Le potentiel hydrique dans la plante et le potentiel de base
Les grandeurs précédentes, qui expriment des quantités d’eau dans les organes d’une plante, sont largement utilisées par les chercheurs, ainsi que par les agronomes et les forestiers. Ces grandeurs ne sont cependant pas suffisantes pour caractériser l’état hydrique des plantes, qui est l’énergie de liaison de l’eau avec un tissu ou un organe. En effet, l’humidité ou la teneur en eau relative ne permettent de déterminer le sens ni l’intensité des flux d’eau, par exemple entre différentes parties d’une plante, ou bien entre le sol et les racines. On peut s’en convaincre avec cet exemple : un sol, même bien humide, n’a pas en général une humidité pondérale supérieure à 1, alors que celle des racines est supérieure à 5, parfois beaucoup plus (Tableau 1) ; pourtant l’eau circule du sol vers les racines.
Rappels
Il est établi depuis longtemps que le potentiel hydrique, souvent noté Y (voir chapitres I.2 et II.6) détermine le sens et l’intensité des mouvements de l’eau le long du trajet sol-plante-atmosphère. Nous donnons ci-dessous quelques rappels sur les notions les plus importantes, le lecteur pouvant se référer aux chapitres qui détaillent ces notions :
· le potentiel hydrique d’un tissu végétal ou d’un échantillon de sol est exprimé par rapport à un état de référence, celui de l’eau pure (à la même température que celle de l’échantillon et à la pression atmosphérique) pour laquelle par définition Y = 0 ;
· par convention, Y est positif si l’eau de l’échantillon cède spontanément de l’énergie pour revenir à l’état de référence (l’eau est alors sous pression), ou au contraire négatif s’il faut lui fournir de l’énergie, par exemple une pression, pour revenir à cet état (l’eau est sous tension) ;
· en conditions naturelles, le potentiel hydrique des différentes parties de la plante est le plus souvent négatif : l’eau est « liée » à la plante. Il peut néanmoins être positif dans certaines conditions, notamment au printemps chez certaines espèces (bouleau, noyer, vigne) ;
· dans la plante, Y est généralement apporximé par la somme de deux composantes, le potentiel de turgescence (Y P) et le potentiel osmotique (Y p) : Y = Y P + Y p ; noter que Y P est positif et Y p est négatif ;
· enfin, les mouvements de l’eau se font dans le sens des potentiels les plus élevés vers les plus faibles. Un modèle simple, l’analogie électrique, qui est calquée sur la célèbre loi d’Ohm, permet de relier le flux d’eau (F) sur un trajet donné, une tige ou une racine par exemple, à la différence des potentiels entre ses deux extrémités (Y e et Y s, où les lettres e et s désignent l’entrée et la sortie de ce trajet). Cette loi simple s’écrit : Y e - Y s = Rh . F, dans laquelle Rh désigne la résistance hydraulique au transfert de l’eau. Cette notion est bien connue des plombiers qui savent que lorsqu’on diminue la résistance hydraulique des canalisations, en utilisant des diamètres de tuyaux plus importants, on augmente le débit de l’eau au robinet ! Ainsi, le potentiel hydrique dans les différents organes de la plante évolue-t-il en parallèle avec sa transpiration ; on ne peut pas traiter indépendamment la variation de chacune de ces deux grandeurs.
Le potentiel hydrique de la plante enracinée
Quantifier la disponibilité en eau de la zone de sol dans laquelle la plante est enracinée, en d’autres termes son potentiel hydrique (Ys) est particulièrement important pour en expliquer les réactions physiologiques, la croissance, voire le dépérissement et la mortalité. Malheureusement, la mesure du Ys est très souvent problématique, surtout en condition de sécheresse. En effet, lorsque la plante transpire, on observe que les couches de sol superficielles se dessèchent plus rapidement que les couches les plus profondes, notamment en raison de la plus forte densité racinaire à ces niveaux. Les chercheurs ont pu montrer qu’il était possible de contourner cette difficulté. Le potentiel hydrique de la plante varie dans la journée, de façon sensiblement synchrone avec sa transpiration. Il diminue dans la journée jusqu’aux alentours de midi solaire (14 heures à l’horloge, en France, en été). Le potentiel augmente ensuite progressivement le soir et la nuit pour plafonner à une valeur maximale et stable, alors que la transpiration a cessé depuis plusieurs heures. En effet, lors de la plupart des nuits, l’atmosphère, qui s’est refroidie, se trouve à un niveau d’humidité relative proche de la saturation ; parallèlement, le rayonnement est nul ou quasi-nul. La valeur du potentiel hydrique à ce moment-clé est appelée potentiel de base (Yb) et constitue une donnée écophysiologique de référence : lorsque la plante atteint Yb, les potentiels hydriques sont égaux dans toute la plante (cf. chapitre II.6). Ceci est vrai au gradient de potentiel hydrostatique près, qui est de 0,01 MPa (=0,1 bar) par mètre de hauteur. En conséquence, mesurer le potentiel de base de la plante revient à mesurer le potentiel hydrique de son système racinaire à l’interface avec le sol. En d’autres termes, Yb représente le potentiel hydrique du sol « vu » par la plante. Cette mesure est généralement réalisée avec une chambre à pression, qui sera décrite plus loin.
2.3. La capacitance hydraulique
Quand un échantillon perd ou gagne de l’eau, deux grandeurs varient simultanément, son contenu en eau et son potentiel hydrique. On peut mettre en évidence, pour un organe ou un tissu, une relation entre ces deux grandeurs. Cette relation s’appelle la capacitance hydraulique Ch. Elle s’exprime comme le rapport entre une variation de volume d’eau et la variation de son potentiel hydrique qui la provoque, soit :
Ch = dVe/dY (6)
avec :
- dVe : variation de la quantité d’eau dans l’échantillon (exprimée par unité de masse ou de volume) ;
- dY : variation de potentiel hydrique (en Pa ou MPa).
Ch s’exprimera donc par exemple en cm3 par MPa ou en g par MPa.
Cette définition de la capacitance hydraulique est rigoureusement équivalente à celle de la capacité électrique d’un condensateur, qui est définie comme le rapport entre la quantité d’électricité, Q, portée par ses plaques et la différence de potentiel auxquelles sont soumises celles-ci. La formule est en électricité : C = dQ/dV.
Une similitude simple, celle de l’éponge humide que l’on presse, permet de saisir intuitivement la signification physique de la capacitance. Si on plonge l’éponge dans une bassine remplie d’eau, elle va s’imbiber d’eau. Si on l’en sort, toute l’eau qui ne peut être retenue dans l’éponge (et qui correspond à un Y légèrement positif) va s’écouler. Quand l’éponge ne gouttera plus, elle sera saturée en eau : le ψ de l’eau de l’éponge et celui de l’eau de la bassine auront alors pratiquement la même valeur, 0. Une fois dans cet état, si l’on veut faire sortir une certaine quantité d’eau de l’éponge, il faut exercer avec les mains une certaine pression, c’est-à-dire fournir un certain travail. Il y a ainsi une relation étroite et mesurable entre la quantité d’eau extraite et la valeur de la pression exercée. On peut continuer ce protocole très simple et on verra que moins l’éponge contient d’eau, plus il faudra fournir de travail pour extraire une même quantité d’eau. Il en est de même pour un sol ou pour un échantillon végétal. Cela signifie que la capacitance hydraulique d’un échantillon végétal n’est pas une constante, mais varie avec le potentiel hydrique de l’échantillon, contrairement à la capacité d’un condensateur électrique. La plupart des tissus de la plante cèdent facilement de l’eau lorsque leur état d’hydratation est élevé et, a contrario, en cèdent difficilement lorsque le tissu est desséché, comme c’est le cas dans l’exemple de l’éponge.
Capacitance hydraulique et réserves hydriques
Ces deux expressions sont liées, mais non équivalentes. En effet, si la capacitance hydraulique d’un organe représente la quantité d’eau perdue lorsqu’on diminue le potentiel hydrique d’une certaine valeur, la réserve hydrique est tout simplement la quantité d’eau totale que peut perdre un organe ou une plante en partant de son état maximum d’hydratation. Noter qu’il faut distinguer entre la réserve hydrique totale et celle qui est réversible, qui peut donc se reconstituer lorsque la plante est remise dans des conditions favorables d’alimentation en eau.


