Conclusion et annexes
Conclusion
Nous avons vu que la réponse d’une plante aux facteurs de son environnement dépendait de paramètres et variables-clés, relativement moins nombreux pour ce qui est de son fonctionnement hydrique que pour son fonctionnement carboné.
La modélisation permet de reproduire les flux, potentiels et teneurs en eau dans la plante. Nous avons vu aussi qu’en faisant varier les paramètres qui caractérisent une espèce et les variables de l’environnement (sol, climat), on mettait en évidence des comportements contrastés, entre des plantes économes en eau et d’autre plus dépensières. Les paramètres majeurs caractérisant la plante sont morphologiques (surfaces foliaire et racinaire, tissus conducteurs, volume des tissus échangeurs d’eau…) et physiologiques (régulation stomatique, vulnérabilité à la cavitation…). Il ne faut toutefois pas penser que le modélisateur peut utiliser toutes les combinaisons de ces paramètres, car la plante montre des ajustements, résultant de son évolution. Un exemple d’ajustement dynamique est la coordination généralement observée chez les plantes supérieures entre l’abaissement de leur potentiel hydrique et la fermeture de leurs stomates, permettant de limiter la dégradation de leur xylème lorsque la sécheresse devient trop intense.
Si la plupart des exemples présentés ci-dessus comparent des types de fonctionnement volontairement très contrastés, on trouve chez les plantes terrestres tous les comportements intermédiaires. Chaque espèce a trouvé un optimum de fonctionnement, adapté à son milieu. L’amélioration génétique peut malgré tout accentuer un caractère recherché par l’agriculteur ou le forestier, mais le plus souvent il sera nécessaire d’apporter des modifications à l’environnement de la culture ou de l’arbre : irrigation, fertilisation, gestion de la densité de culture, des pathogènes…
Il existe un système d’unités international (SI) utilisé par les physiciens, les chimistes et adopté par les biologistes.
Nous donnons ci-dessous les unités dans lesquelles s’expriment les différentes variables et paramètres que nous utilisons dans le modèle. Toutefois, d’autres unités sont employées, notamment parce que le système SI conduit à des ordres de grandeurs trop grands ou trop petits.
|
Variable ou paramètre |
abréviation |
Unité SI |
autres unités |
|
transpiration |
T |
Kg.m-2.s-1 |
mm.h-1 |
|
évapotranspiration potentielle |
ETP |
||
|
potentiel hydrique du sol |
Ys |
Pa ou MPa |
bar |
|
potentiel hydrique de la feuille |
Yf |
||
|
potentiel hydrique du xylème |
Yx |
||
|
teneur en eau relative |
TER |
sans unité |
|
|
conductance hydraulique totale (des racines aux feuilles) |
Kh |
Kg.m-2.s-1.Pa-1 |
mm.h-1.MPa-1 |
Annexe 2. Expression de la formule de Penman
La formule de Penman peut s’écrire comme la somme de deux termes : un terme radiatif (rad) et un terme convectif (conv). Le chapitre II.5 présente en détail cette formule et sa démonstration physique.
ETP est exprimée ici en mm d’eau, 1 mm d’eau correspondant à 1 kg d’eau par m² :
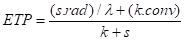
avec :
rad = Rg . (1 - a) – Ri
conv = 0,26 . D. (1 + 0,54 . V)
Ri : rayonnement de grande longueur d’onde, émis par la végétation vers l’atmosphère. Ri est calculé à partir de la température de la surface végétale, souvent assimilée à celle de l’air (T) ;
s : dérivée de la courbe de vapeur saturante (en Pa.°C-1)
Variables météorologiques mesurées :
Rg : rayonnement global (en MJ.m-2)
V : vitesse du vent (en m.s-1)
D : déficit de saturation de l’air (en Pa)
T : température de l’air (en °C)
Constantes :
a : albédo. C’est la fraction du rayonnement qui est réfléchie vers l’atmosphère ; pour la plupart des végétations, a est compris entre 0,1 et 0,2
l : chaleur latente de vaporisation de l’eau (= 2,45 106 J.kg-1)
k : constante psychrométrique (= 66 Pa.°C-1)
Annexe 3 : Une fonction mathématique polyvalente pour représenter la réponse des plantes aux variations des facteurs du milieu
En écophysiologie, nous utilisons souvent des fonctions qui décrivent la réponse aux forçages ou aux contraintes, aussi bien d’origine climatique (température, rayonnement…) qu’édaphique (sécheresse, excès d’eau…). Beaucoup de phénomènes qui régissent la réponse de la plante (les échanges d’eau, de dioxyde de carbone, la croissance, etc.) ne sont pas linéaires.
Certaines réponses à une contrainte sont fortes dès le développement de la contrainte, mais, au fur et à mesure que cette dernière augmente, la réponse de la plante se ralentit. La Fig. MOD14A en donne une illustration, ici l’exemple de la photosynthèse d’une feuille en réponse à l’augmentation de l’énergie lumineuse reçue. En revanche, d’autres contraintes sont peu suivies d’effet à leur début et ce n’est que lorsqu’elles deviennent suffisamment fortes que la plante réagit de façon importante. C’est l’exemple de la croissance en réponse au dessèchement du sol montré sur la Fig. MOD14B.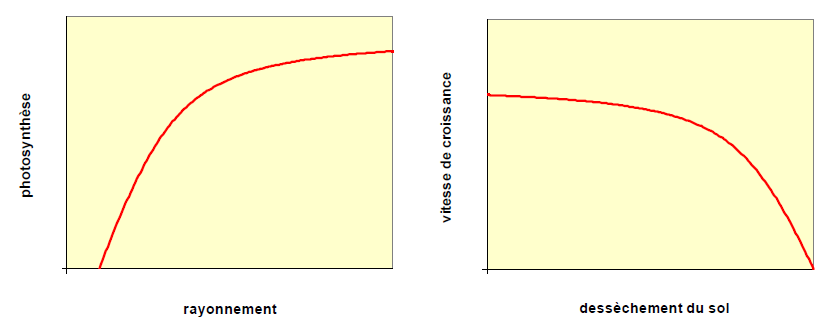
Figure MOD14. Exemple de deux types de réponse à une contrainte ou un forçage exercé par le milieu environnant. En A, photosynthèse d’une feuille en fonction du rayonnement reçu. En B : effet du dessèchement du sol sur l’accroissement en biomasse d’une plante.
Nous utilisons dans le modèle des fonctions non linéaires appelées hyperboles non équilatères, qui s’écrivent :
![]() (équation 5)
(équation 5)
avec :
x : la variable dépendante, dite de forçage (ETP, par exemple) ;
a, b, c : les paramètres de la fonction
D’autres fonctions, plus ou moins compliquées sont évidemment utilisées.
Bibliographie
Granier A, Bréda N, Biron P, Villette S (1999) A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecological Modelling, 116, 269-283.
Granier A, Bréda N, Claustre, JP, Colin F (1989) Variation of hydraulic conductance of some adult conifers under natural conditions. In Annales des Sciences Forestières, Vol. 46, No. Supplement, pp. 357s-360s.
Tuzet A, Perrier A, Leuning R (2003) A coupled model of stomatal conductance, photosynthesis and transpiration. Plant, Cell & Environment, 26(7), 1097-1116.
Tuzet A, Granier A, Betsch P, Peiffer M, Perrier A (2017) Modelling hydraulic functioning of an adult beech stand under non-limiting soil water and severe drought condition. Ecological Modelling, sous presse.
Sommaire :


